
Задание 1
Приведение показателей в сопоставимый вид предполагает:
1) Нахождение абсолютного отклонения фактического значения показателя от планового или базисного.
2) Нейтрализацию воздействия различных факторов путем приведения анализируемых показателей к единому базису.
3) Нахождение относительного отклонения фактического значения показателя от планового или базисного.
4) Деление изучаемой совокупности объектов на качественно однородные группы.
Задание 2
Ассортимент продукции это…
1. перечень наименований изделий
2. соотношение отдельных изделий к общему объему продукции
3. перечень изделий с определением количества каждого из них
4. соотношение отдельных видов продукции между собой
Задание 3
Отношение стоимости новых основных средств к стоимости основных средств на конец периода это коэффициент
Ответ:
поступления
выбытия
обновления
ликвидации
Задание 4
Наличие материальных ресурсов на предприятии отражает:
1. выход отходов
2. материалоемкость
3. коэффициент выхода годного продукта
4. остатки материалов на складе
Задание 5
Темп роста выручки от продажи продукции составил 106,18%, темп роста среднесписочной численности - 104,18%. Эффективность использования трудовых ресурсов:
1) увеличилась;
2) снизилась;
3) осталась неизменной.
Задание 6
На структуру себестоимости продукции влияют следующие факторы:
1) Величина затрат на один рубль продукции
2) Специфика отрасли, географические и природно-климатические условия, технико-организационный уровень предприятия.
3) Уровень переменных затрат на единицу продукции и сумма постоянных затрат на весь объем выпуска продукции.
4) Уровень прибыли и цены на продукцию.
Задание 7
По данным финансовой отчетности, общая рентабельность капитала организации рассчитывается как отношение:
1) прибыли от продаж к средней стоимости капитала;
2) прибыли до налогообложения к средней стоимости собственного капитала;
3) прибыли до налогообложения к средней стоимости капитала
Задание 8
Собственный капитал предприятия по балансу определяется как:
1. итог III раздела "Капитал и резервы"
2. итог III раздела + итог раздела IV
3. уставный капитал
Задание 9
Коэффициент утраты платежеспособности рассчитывается для организаций, у которых:
1) коэффициент текущей ликвидности и коэффициент обеспеченности оборотных активов собственными оборотными средствами соответствуют нормативным ограничениям
2) коэффициент текущей ликвидности не соответствует нормативному ограничению
3) либо коэффициент текущей ликвидности, либо коэффициент обеспеченности оборотных активов собственными оборотными средствами, либо оба вышеназванных коэффициента не соответствуют нормативным ограничениям.
Задание10
Для определения типа финансовой устойчивости следует сравнить:
1.собственный капитал с заемным капиталом
2.оборотные активы с краткосрочными обязательствами
3.запасы и затраты с источниками их покрытия
Задание 11
Укажите экстенсивный фактор развития производства:
1) увеличение количества использованных ресурсов, увеличение времени использования ресурсов;
2) совершенствование качественных характеристик использования ресурсов
Задание 12
Под сезонностью спроса понимают:
1) степень сохранения спроса на товар в течение годовых сезонов;
2) устойчивую закономерность внутригодичной динамики изменяющегося в течение года покупательского спроса на соответствующие товары в связи с изменением сезонов года;
3) связанные со сменой времени года затраты по хранению товаров.
Задание 13
Техническое состояние основных фондов анализируется с помощью показателей:
Ответ:
поступления, выбытия
годности, износа
фондоотдачи, фондоемкости
Задание 14
К обобщающим показателям эффективности использования материальных ресурсов НЕ относятся:
1.материалоемкость
2.сырьеемкость
3.доля материальных затрат в себестоимости
Задание 15
Темпы роста среднегодовой выработки рабочего ниже, чем среднедневной. Причина различий динамики этих показателей связана с:
1. целодневными потерями рабочего времени
2. внутрисменными потерями рабочего времени
3. ухудшением структуры персонала
Задание 16
Если фактический выпуск продукции ниже запланированного можно ожидать, что будут ниже заявленных в смете следующие виды затрат:
1) общие переменные издержки
2) общие постоянные издержки
3) переменные издержки на единицу продукции
4) постоянные издержки на единицу продукции
Задание 17
Для определения размера чистой прибыли, приходящейся на один рубль собственного капитала, используется показатель:
1) коэффициент оборачиваемости собственного капитала;
2) коэффициент рентабельности собственного капитала,
3) коэффициент маневренности собственного капитала.
Задание 18
Собственный оборотный капитал определяется:
1. оборотные активы - собственный капитал
2. оборотные активы + текущие обязательства
3. собственный капитал - внеоборотные активы
Задание 19
Перспективы платежных возможностей характеризует коэффициент:
1. обеспеченности собственными средствами
2. текущей ликвидности
3. абсолютной ликвидности
Задание 20
Если общая величина запасов полностью покрывается с учетом привлечения собственных оборотных средств и долгосрочных кредитов, то имеет место:
1) нормальная устойчивость;
2) неустойчивое состояние;
3) абсолютная устойчивость.
4) кризисное состояние.
Задание 21
Относительная экономия (перерасход) отдельных видов производственных ресурсов характеризует изменение:
1. объема выпущенной продукции
2. эффективности использования ресурсов
3. расхода ресурсов на производство
Задание 22
При анализе качества продукции НЕ выделяют показатели:
Ответ:
обобщающие
индивидуальные
косвенные
частные
Задание 23
Движение основных средств анализируется с помощью показателей:
1. поступления, выбытия, обновления
2. годности, износа
3. фондоотдачи
Задание 24
Выберите наиболее правильный ответ. Общая сумма прямых материальных затрат зависит от:
1) Объема продаж и структуры продукции.
2) Уровня материальных затрат на единицу продукции.
3) Объема выпуска продукции, структуры продукции, уровня материальных затрат на единицу продукции
4) Объема выпуска продукции, структуры продукции, уровня материальных и трудовых затрат на единицу продукции
Задание 25
Приняты следующие обозначения:
ГВ - среднегодовая выработка рабочих, руб.
ЧВ - среднечасовая выработка рабочих, руб.
Г - Продолжительность рабочего года, дн.
Д - продолжительность рабочего дня, час.
Для анализа годовой выработки рабочих может использоваться формула:
1) ГВ = Г · Д · ЧВ
2) ГВ = Г · ЧВ
3) ГВ = (Г / Д) · ЧВ
Задание 26
Рост переменных затрат может происходить в результате:
1.роста производства продукции
2.изменения организационной структуры
Задание 27
Маржинальный доход предприятия равен:
1. максимальной прибыли
2. сумме прибыли и переменных затрат
3. разница между выручкой от продаж и прямыми переменными затратами
Задание 28
Анализ имущественного состояния хозяйствующего субъекта.
1) оценка стоимости его активов, изучение структуры имущества ее динамики
2) оценка структуры пассивов, соотношения собственных и заемных средств, вложенных в имущество
3) соотношение стоимости активов и структуры пассивов организации
Задание 29
Коэффициент утраты платежеспособности рассчитывается за период равный ...
Ответ:
шести месяцам
трем месяцам
одному месяцу
Задание 30
При анализе абсолютных показателей финансовой устойчивости НЕ выделяют тип финансовой устойчивости
Ответ:
неустойчивое состояние
нормальная устойчивость
ненормальная устойчивость
Задание 31
Для оценки структуры показателей рассчитывают
Ответ:
темп роста
абсолютное отклонение
удельный вес
Задание 32
Обобщающим показателем качества продукции является:
1. полезность продукции
2. удельный вес сертифицированной продукции
3. удельный вес бракованной продукции
Задание 33
Структуру основных средств характеризует:
Ответ:
доля активной части ОПФ
коэффициент обновления
коэффициент выбытия
коэффициент роста
Задание 34
При анализе обеспеченности организации материальными ресурсами оценку обычно проводят по
1.всем видам ресурсов
2.занимающим наибольшую долю в материальных затратах
Задание 35
При анализе показателей производительности труда, если темпы роста часовой выработки рабочих отстают от темпов роста дневной выработки рабочих. Это означает, что имело(и) место:
1) ухудшение структуры персонала;
2) целодневные потери рабочего времени;
3) внутрисменные потери рабочего времени.
Задание 36
Сумма переменных затрат в связи с ростом объема продукции
1. не изменится
2. увеличится пропорционально росту продукции
3.увеличится не пропорционально росту продукции
Задание 37
Увеличению рентабельности активов способствует рост:
1) стоимости активов;
2) прибыли до налогообложения;
3) выручки от продаж;
Задание 38
Для оценки структуры источников средств организации необходимо использовать показатели:
1) капитализации;
2) ликвидности;
3) верно 1 и 2.
Задание 39
Оптимальное значение коэффициента абсолютной ликвидности составляет
1) ≤ 0,2
2) ≤ 2,0
3) ≥ 0,2
4) ≥ 2,0
Задание 40
Ускорение оборачиваемости активов способствует:
1) росту рентабельности активов
2) уменьшению рентабельности активов
3) росту рентабельности продукции
Задание 41
Для оценки степени использования достижений науки и поиска резервов осуществляют сравнение достигнутых показателей:
1. со средними показателями по зоне области
2. анализируемого предприятия с лучшими результатами
3. с нормативными показателями
Задание 42
Внутренние причины аритмичности:
1. несвоевременная поставка сырья и материалов поставщиками
2. низкий уровень организации производства
3. тяжелое финансовое состояние предприятия
Задание 43
Коэффициент обновления основных средств определяется по формуле:
a) Стоимость вновь введенных в эксплуатацию средств / Общая стоимость основного капитала на конец года
2) Стоимость вновь введенных в эксплуатацию средств / Общая стоимость основного капитала на начало года
3) Стоимость вновь введенных в эксплуатацию средств / Стоимость имущества
Задание 44
При анализе эффективности использования материальных ресурсов положительно оценивают:
1) рост материалоемкости
2) снижение материалоемкости
3) рост стоимости материальных ресурсов
4) снижение материалоотдачи
Задание 45
Приняты обозначения:
О - общее число отработанных человеко-часов работниками
Ч - среднесписочная численность работников
Г - продолжительность рабочего года
Д - Продолжительность рабочего дня
Тд - общее число отработанных человеко-дней работниками
Формула для расчета влияния факторов на общее число отработанных человеко-часов работниками имеет вид:
1. О = Ч * Г * Д * Тд
2. О = Ч * Тд * Д
3. О = Ч * Г * Д
Задание 46
По способу отнесения на себестоимость затраты делятся на:
1. постоянные и переменные
2. прямые и косвенные
3. контролируемые и неконтролируемые
Задание 47
Рентабельность продаж рассчитывается как:
1) средняя стоимость собственных средств / выручка от продаж
2) прибыль от продаж / выручка от продаж
3) стоимость имущества / выручка от продаж
Задание 48
В пассиве баланса НЕТ раздела:
Ответ:
капитал и резервы
собственный капитал
долгосрочные обязательства
краткосрочные обязательства
Задание № 49
Коэффициент восстановления платежеспособности рассчитывается за период равный ...
Ответ:
шести месяцам
трем месяцам
одному месяцу
Задание 50
Долю собственного капитала, направленного в оборотные активы характеризует коэффициент
Ответ:
маневренности
автономии
структуры имущества
Задание 51
Выход готовой продукции на 1 руб. ресурсов характеризуют показатели:
Ответ:
емкости
отдачи
Задание 52
Для оценки уровня механизации производства используют следующий показатель:
1) степень охвата рабочих механизированным трудом;
2) коэффициент годности оборудования;
3) уровень механизированного труда в общих трудозатратах;
Задание 53
Темпы роста фондоотдачи активной части основных средств выше, чем темпы фондоотдачи всех производственных фондов. Причина различий темпов роста этих показателей отражает:
1.рост доли активной части
2.снижение доли активной части
3.улучшение использования активной части ОПФ
Задание 54
На объем выпуска продукции, по факторам, связанным с предметами труда прямо НЕ влияют:
1. стоимость материальных затрат
2. остатки материалов на складе
3. материалоемкость
Задание 55
При росте производительности труда и прочих равных условиях:
1) объем выпуска продукции уменьшается;
2) объем выпуска продукции не меняется;
3) объем выпуска продукции увеличивается.
Задание 56
Темпы роста оплаты труда должны:
1) Быть равны темпам роста производительности труда
2) Превышать темпы роста производительности труда
3) Быть меньше, чем темпы роста производительности труда
рат на 1 рубль продукции и уровень инфляции
2. уровень инфляции и качество продукции
3. качество продукции и объем продаж
4. объем продаж и уровень затрат на 1 рубль продукции
Задание 58
Величину собственного оборотного капитала по балансу можно рассчитать:
1.внеоборотные активы - собственный капитал
2.собственный капитал - внеоборотные активы
3.собственный капитал + долгосрочные пассивы
Задание 59
Ликвидность баланса отражает:
1) состояние имущества и обязательств, при котором организация подлежит ликвидации
2) способность организации покрывать обязательства активами, срок превращения которых в денежную форму соответствует сроку погашения обязательств
3) период утраты платежеспособности организации
Задание 60
Эффективность использования имущества характеризует коэффициент
1.Рентабельность собственного капитала
2.Оборачиваемость собственного капитала
3.Оборачиваемости имущества
Задание 61
Горизонтальный метод финансового анализа - это:
1) сравнение каждой позиции отчетности с предыдущим периодом
2) определение структуры итоговых финансовых показателей
3) определение основных финансовых коэффициентов
Задание 62
Индивидуальными показателями качества продукции являются показатели, характеризующие:
1. одно из ее свойств
2. потери от брака
3. уровень модернизации
Задание 63
Уровень стоимости основных производственных средств на единицу продукции определяют с помощью показателя:
1) фондоотдачи;
2) фондоемкости;
3) фондовооруженности.
Задание 64
Выберите наиболее правильный ответ. Причиной роста удельного веса производственных запасов могут быть:
1) затоваривание, падение сбыта продукции;
2) желание руководства организации произвести ликвидные вложения;
3) увеличение объема нематериальных активов.
Задание 65
Показатель производительности труда показывает стоимость основных средств в расчете на одного работника предприятия
Ответ:
не верно
верно
Задание 66
Объектами анализа себестоимости продукции НЕ являются:
1. полная себестоимость товарной продукции
2. затраты на рубль товарной продукции
3. выявление резервов трудовых затрат
4. себестоимость отдельных явлений
Задание 67
К внутренним факторам изменения прибыли НЕ относятся:
1) номенклатура затрат;
2) нарушения к требованию качества продукции;
3) цены на производственные ресурсы;
4) объемы продаж.
Задание 68
Между величиной собственного оборотного капитала (СОК) и суммой оборотных активов НЕ может быть соотношения:
Ответ:
СОК < оборотные активы
СОК > оборотные активы
СОК = оборотные активы
Задание 69
Коэффициент абсолютной ликвидности рассчитывается:
1) отношение собственных средств к валюте баланса
2) отношение наиболее ликвидных активов к сумме наиболее срочных обязательств
3) частное от деления величины денежных средств, краткосрочных ценных бумаг и расчетов на сумму краткосрочных обязательств организаций
Задание 70
Продолжительность оборота имущества (в днях) отражает коэффициент:
1) Выручка от продаж / Валюта баланса
2) Выручка от продаж / Оборотные активы
3) Число дней в периоде / Коэффициент оборачиваемости имущества
4) Коэффициент оборачиваемости имущества / Число дней в периоде
Задание 71
Цепной индекс.
1) характеризует изменение показателя данного периода по сравнению с показателем предыдущего периода
2) отражает изменение показателя данного периода по сравнению с показателем периода, принятого за базу для сравнения
3) равен произведению базисных индексов
Задание 72
Какой показатель анализа качества продукции НЕ относится к группе косвенных показателей:
1.доля модернизированной продукции
2.доля брака в себестоимости
3.штрафы за некачественную продукцию
Задание 73
К факторам экстенсивной загрузки основных средств НЕ относят:
1. использование времени
2. производительность единицы оборудования
Задание 74
Показатель, характеризующий долю материальных затрат в выручке от продаж, показывает:
1) трудоемкость продукции;
2) материалоемкость продукции;
3) отдачу от материальных ресурсов.
Задание 75
При анализе показателей производительности труда, если темпы роста годовой выработки рабочих отстают от темпов роста производительности труда всего промышленно-производственного персонала. Это означает, что имело(и) место:
1) ухудшение структуры персонала;
2) целодневные потери рабочего времени;
3) внутрисменные потери рабочего времени.
Задание 76
Метод калькулирования себестоимости применяемый в единичном и мелкосерийном производстве...
Ответ:
попроцессный
нормативный
позаказный
Задание 77
При факторном анализе прибыли от продаж НЕ рассчитывают влияние фактора:
1) структуры и ассортимента продукции
2) проценты к уплате;
3) себестоимость продукции;
4) цену единицы продукции
Задание 78
Пассив баланса содержит сведения о
Ответ:
размещении капитала
источниках средств
финансовой устойчивости
уровне ликвидности
Задание 79
Отношение всех текущих активов к краткосрочным обязательствам называется коэффициентом:
Ответ:
текущей ликвидности
абсолютной ликвидности
срочной ликвидности
Задание 80
Оптимальное значение коэффициента соотношения заемного и собственного капитала составляет
1) ≤ 1,0
2) ≤ 2,0
3) ≥ 1,0
4) ≥ 2,0
Задание 81
По способу выявления резервы делят на:
Ответ:
явные и скрытые
текущие и перспективные
интенсивные и экстенсивные
Задание 82
Коэффициент ритмичности продаж определяется отношением:
1) суммы фактического объема продаж за исследуемый период к сумме планового объема продаж;
2) суммы фактического объема продаж не выше плана за исследуемый период к сумме планового объема продаж;
3) суммы фактического объема продаж за исследуемый период к сумме затрат.
Задание 83
Уровень затрат основных производственных фондов на единицу продукции определяют с помощью показателя:
Ответ:
фондоотдачи
фондоемкости
фондовооруженности
Задание 84
Выберите наиболее правильный ответ.
Показателями эффективного использования материальных ресурсов считается:
1) Повышение материалоемкости, материалоотдачи, рост прибыли на 1 рубль материальных затрат.
2) Повышение материалоемкости, снижение материалоотдачи, снижение прибыли на 1 рубль материальных затрат.
3) Снижение материалоемкости, повышение материалоотдачи, рост прибыли на 1 рубль материальных затрат.
4) Снижение материалоемкости, снижение материалоотдачи, рост прибыли на 1 рубль материальных затрат.
Задание 85
К обобщающим показателям производительности труда относятся:
1) Технологическая трудоемкость.
2) Среднегодовая, среднедневная, среднечасовая выработка продукции одним рабочим (основным работником), среднегодовая выработка на одного работающего в стоимостном выражении.
3) Трудоемкость обслуживания производства и трудоемкость управления.
4) Среднечасовая выработка рабочего.
Задание 86
Рост постоянных затрат может происходить в результате:
1.повышения цен на реализуемую продукцию
2.увеличения натурального объема реализации
3.повышения оплаты труда работников
Задание 87
Прибыль от продаж рассчитывается как
1) Разница между выручкой от продаж и себестоимостью проданных товаров, работ, услуг
2) Разница между выручкой от продаж себестоимостью проданных товаров (работ, услуг), управленческими и коммерческими расходами
3) Разница между фактическим и безубыточным объемом продаж
Задание 88
Какие статьи НЕ относятся к Капиталу и резервам
Ответ:
Уставный капитал
Кредиторская задолженность
Нераспределенная прибыль
Добавочный капитал
Задание 89
Если в динамике темпы прироста оборотных активов 25%, а внеоборотных активов - 18 %, то это свидетельствует о тенденции:
1) ускорения оборачиваемости всех активов;
2) замедления оборачиваемости активов;
3) роста собственных оборотных средств.
Задание 90
Оптимальное значение коэффициента обеспеченности запасов источниками их формирования составляет
1) ≤ 1,0
2) ≤ 0,8
3) ≥ 1,0
4) ≥ 0,6
Задание 91
Какой показатель рассчитывается при горизонтальном анализе
Ответ:
удельного веса
структуры
темпа роста
Задание 92
Отношение стоимости сертифицированной продукции к общему объему выпуска продукции является показателем для оценки:
1) ассортимента продукции;
2) ритмичности;
3) качества продукции;
4) эффективности использования материальных ресурсов.
Задание 93
Экстенсивное использование основных производственных фондов характеризует:
Ответ:
фондоемкость, фондоотдача
коэффициент сменности
фондовооруженность труда
Задание 94
Отношение объема выпуска продукции к стоимости материальных затрат является показателем для оценки:
1) себестоимости продукции;
2) ритмичности;
3) качества продукции;
4) эффективности использования материальных ресурсов
Задание 95
Трудоемкость продукции определяется как отношение:
1) Среднегодовая величина основных средств / Среднесписочная численность работников
2) Среднесписочная численность работников / Выпуск продукции
3) Выпуск продукции / Среднегодовая величина основных средств
Задание 96
Приняты следующие обозначения
ФОТ - фонд оплаты труда
Ч - численность персонала
ЗП - среднегодовая заработная плата
Влияние изменение численности на фонд оплаты труда рассчитывается по формуле
1.(Ч1 - Ч0) х ЗП0
2.(Ч0 - Ч1) х ЗП0
3.(ЗП1 - ЗП0) х Ч1
4.(ЗП1 - ЗП0) х Ч0
Задание 97
Приняты следующие обозначения:
Рпр - рентабельность реализованной продукции; Кф\емк - коэффициент фондоемкости; Кзакр - коэффициент закрепления.
Влияние коэффициента закрепления на уровень общей рентабельности рассчитывается
1. Рпр 1 /(КФ/емк1 + Кзакр0) - Рпр 1/(КФ/емк0 + Кзакр0)
2. Рпр 1 /(КФ/емк0 + Кзакр0) - Рпр 1/(КФ/емк1 + Кзакр0)
3. Рпр 0 /(КФ/емк1 + Кзакр0) - Рпр 1/(КФ/емк0 + Кзакр0)
4. Рпр 1 /(КФ/емк1 + Кзакр1) - Рпр 1/(КФ/емк1 + Кзакр0)
Задание 98
Мобильные активы находятся в разделе
Ответ:
Внеоборотные активы
Оборотные активы
Капитал и резервы
Краткосрочные обязательства
Задание 99
Наиболее ликвидная часть оборотных средств предприятия:
Ответ:
незавершенное производство
дебиторская задолженность
сырье и материалы
денежные средства
Задание 100
Как определяется период погашения (в днях) кредиторской задолженности :
1) себестоимость проданных товаров / средние остатки кредиторской задолженности;
2) стоимость кредиторской задолженности на конец года / выручка от продажи;
3) 365 / коэффициент оборачиваемости кредиторской задолженности (в оборотах).
�������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������ное пространственно-временное состояние организма в каждой области многомерной структуры высших биосистем, где в данный момент находится бризер. Наличие тепловых возмущений (kT) молекулы ДНК, а также возможность существования фуранозных колец нуклеотидов в виде двух конформаций, приводят к усложнению модели и необходимости введения в нее фазовых флуктуаций электронной плотности.
Однако, учитывая, что спектр ФПУ может служить преобразователем стохастических колебаний в детерминированные, стохастическая компонента динамики колебаний электронной плотности в молекуле ДНК является, вероятно, ее атрибутом.
Данная теоретическая модель нелинейной знаковой акусто-электродинамики ДНК легла в основу создания семейства радиоэлектронных устройств - генераторов пакетов уединеных волн в форме возврата Ферми-Паста-Улама (ФПУ-генераторов), предназначенных для генерации электромагнитных волн (солитонов), обладающих характерной пространственно-временной структурой возврата ФПУ, которое выражается в периодическом переходе колебательной структуры от упорядоченного состояния к хаотическому и обратно. При этом в упорядоченном состоянии первоначальная форма волнового пакета и его пространственно-временной спектр полностью повторяются. Важной особенностью ФПУ-генераторов является пространственно-временная структура его поля, которая является относительно простой физической моделью колебательной структуры молекулы ДНК. Это свойство позволяет использовать генератор в экспериментах по исследованию собственных колебаний в препаратах ДНК и по информационному взаимодействию биологических систем, о которых говорилось выше. Первые модели таких генераторов были созданы А. А. Березиным и соавторами (1988, 1989 г. г. ), а затем в 1991г. были принципиально дополнены П. П. Гаряевым и Г. Г. Комиссаровым за счет интеграции в их схемы кодирующего акустического ввода.
Принципиальная схема генератора содержит ФПУ-резонатор в виде двух длинных линий с подключенными к ним нелинейными элементами (туннельные диоды). Напряжение смещения туннельных диодов задается стабилизаторами на транзисторах и стабилитроне. Выбор рабочей точки туннельных диодов и способ их подключения к ФПУ-резонатору обеспечивают форму и спектр колебаний генератора, которые соответствуют нормальным колебаниям одномерной решетки слабо связанных нелинейных (ангармонических) осцилляторов с периодическими граничными условиями, при которых наблюдается явление возврата ФПУ. Для модуляции поля генератора внешними акустическими сигналами может быть использован угольный микрофон. Генератор питается от двух аккумуляторов типа ЦНК 0, 45-I-У2.
С помощью ФПУ-генератора и эмбрионов-доноров удалось непермиссивно дистантно (20 см - 2, 0 м) осуществить эмбриональную индукцию нейральных и мезодермальных производных в ткани эктодермы ранней гаструлы шпорцевой лягушки. Были получены результаты и по восстановлению нативной структуры у аберрантных радиационно поврежденных хромосом пшеницы и ячменя [25,29]. Это показывает реальность существования и моделирования знаковых электромагнитных полей геноволнового уровня, управляющих стратегическим метаболизмом биосистем, их наследственностью, и подтверждает близкие результаты, полученные Дзян Каньджэном.
Существует и другая семантическая ниша знаковых процессов в наследственном аппарате высших биосистем, связанная с его квази-речевыми характеристиками, а также с генетической атрибутикой словообразований в естественных человеческих языках. Ранее получены доказательства, что развитие языков и человеческой речи подчиняется законам формальной генетики. По сути, “тексты” ДНК (квазиречь) и письменность людей, их разговор (истинная речь) выполняют одинаковые управленческие, регуляторные функции, но в разных фрактально-сцепленных масштабированиях. ДНК генетически функционирует на клеточно-тканевом уровне, а человеческая речь, как макрогенетическая структура, используется на уровне общественного суперорганизма. Нам удалось несколько отойти от предшествующей метафоричности использования понятий лингвистики применительно к ДНК, когда произвольно используют термины “слово”, “текст”, “пунктуация”, “грамматика”, интуитивно пытаясь понять иные измерения генома. Такому отходу способствовало применение теории фракталей и метода перекодировок к последовательностям ДНК и структуре текстов людей. Выяснилось, что ДНК и человеческая речь (тексты) обладают стратегически близкой фрактальной структурой в геометрическом смысле. Вероятно, это каким-то образом коррелирует с фрактальной структурой солитонного акустического и электромагнитного ФПУ-поля, генерируемого хромосомным аппаратом высших биосистем. Возможно, именно по этой причине нам удалось зарегистрировать управленческие эффекты на геномах растений, вызываемые с помощью ФПУ-трансформированной человеческой речи, которая резонансно взаимодействует с хромосомной ДНК in vivo [25,29].
Этот результат, осмысленный нами с позиций семиотико-волновой составляющей генетического кода, имеет существенное методологическое значение и для анализа таких суперзнаковых объектов, как тексты ДНК, и для генома в целом. Открываются принципиально новые смысловые ареалы хромосомного аппарата. Однако биологии и гено-лингвистике предстоит пройти еще большой путь, прежде чем картина знаковых рядов ДНК станет относительно ясной и понимаемой. Вводимый нами способ мышления относительно функций генома позволяет сопоставлять различные естественные последовательности ДНК и РНК с оценкой меры их сходства и различия, а также степени относительной сложности их знаковой структуры. И кроме того, что более важно, появляется метод сопоставления смысловых конструкций человеческой речи и кодирующих последовательностей ДНК. Если мы правы в своих логических и экспериментальных построениях, то в общем плане видны новые измерения в понимании мышления и сознания через их отображения в знаковых (смысловых) рядах на разных уровнях организации живой материи - на уровне человеческой речи (высшая форма сознания) и квазиречи генетических молекул (квази-сознание генома). Это хорошо соответствует математико-лингвистической модели Хомского, постулирующей общие принципы, которые лежат в основе любого языка и которые объединяются в “универсальную грамматику”. Такая “универсальная грамматика”, по Хомскому, является врожденной, т. е имеет генетические детерминанты. Это чрезвычайно важное обстоятельство, которое еще раз фокусирует мысль на супергенетическом родстве знаковых структур ДНК и речевых образований человека. В какой-то мере мы подтвердили указанное положение, показав родство фракталей ДНК и человеческой речи. Хомский, вероятно, прав в том, что глубинные синтаксические конструкции, составляющие основу языка, передаются по наследству от поколения к поколению, обеспечивая каждому индивидууму возможность овладеть языком своих предков. То, что ребенок легко учится любому языку, объясняется как раз тем, что в своей основе грамматики всех языков совпадают. Суть человеческого языка инвариантна для всех людей. Можно думать, что эта инвариантность распространяется глубже, достигая макромолекулярных смысловых (“речевых”) структур хромосом. И этому есть определенные теоретико-экспериментальные подтверждения, полученные нами [25,29] и выводящие на существенно значимые методологические подходы мягкого регуляторного вхождения в неизвестные ранее семиотические пласты генетического аппарата высших биосистем. Но в этом же заключена и грозная опасность стратегических семиотико-волновых искажений знакополевого окружения Земли. Идеи волновой (и “речевой”) генетики находятся в фазе активного становления и поэтому необходима система жестко определенных запретов определенных экспериментов в этой области знания, подобная существующей в генной инженерии, например, по клонированию высших организмов.
Независимое подтверждение правильности гипотезы существования квази-вербального или, что одно и то же, образного уровня кодовых функций ДНК (в пределе хромосомного континуума биосистемы) может дать выход из ограниченного, а иногда неверного, функционального поля триплетного генетического кода, не объясняющего ни синтез белковых “текстов”, ни то, как в геноме зашифрована пространственно-временная структура организма. Конечная цель предлагаемого анализа выделение знаковых единиц различных уровней и понимание их семантики в функциональном пространстве ДНК- белок, которое, по крайней мере для ферментов, чрезвычайно гетерогенно (активный центр, сайты узнаваний, архитектоника водородно-гидрофобных сил самоорганизации пептидной цепи). Многоязычный метаболический “разговор” между информационными биополимерами клетки и их функционирование как результат обмена знаковыми биосигналами предполагают два взаимно коррелированных уровня этого обмена - вещественный и волновой. Вещественный хорошо изучен (матричное копирование ДНК-РНК-белки, взаимодействие антиген-антитело, самосборка клеточных структур), а тесно связанный с ним волновой уровень практически не изучен официальной наукой. И ситуация здесь непроста. Электромагнитные и акустические излучения белков, нуклеиновых кислот, мембран и цитоскелета хорошо известны. Представляется, что этот уровень информационных контактов клеточно-тканевого пространства выводит метаболические процессы в полевое измерение со своей “языковой” спецификой и регуляцией.
Рассматриваемые биоинформационные потоки, сцепленные с обменом веществ и энергии, не ограничиваются делением знаковых рядов на вещество и поле, но многократно умножаются фрактальностью этих рядов. Например, в акустико-электромагнитной компоненте сигнальных функций ДНК наблюдается фрактальность солитонного поля, формально описываемого уравнениями в рамках явления возврата Ферми-Паста-Улама. Это еще более усложняет семантический анализ белково-нуклеиновых и иных информационных контактов биоструктур. Можно полагать, что в живых клетках существует иерархия вещественно-волновых знаковых структур, где условную градацию “буква (фонема) - морфема - слово - предложение...” задает фрактальность этих структур. И то, что в одном масштабе является “предложением”, в другом, более крупном, может быть лишь “словом” и т. д. Другая сложность связана с понятием “рамки считывания”. Сдвиг на одну букву (или эквивалентное этому небольшое изменение фазы, поляризации, частоты физических полей в пространстве-времени биосистемы) может полностью поменять смысл читаемого текста (воспринимаемого образа), не говоря уже о том, что сами тексты, к примеру, в одних и тех же последовательностях ДНК могут быть записаны разными языками. Более того, нет запрета на понимание “текстов” жидкокристаллических хромосомных ДНК, как читаемых в трех- или n-мерном пространстве, когда “буквы слов” выстраиваются не только в одну линию и в одном измерении, но “читаются” вдоль и поперек, вверх и вниз и так далее. В таком процессе поочередно создается и уничтожается бесконечный континуум анизотропных “нитей текстов”, идущих во всех направлениях динамичного интерфазного хромосомного континуума всего пространства биосистемы. Предлагаемая логика неизбежна, если мы хотим понять сущность феномена жизни. Сказанное не следует рассматривать как предтечу пересмотра только принятой модели триплетного генетического кода. Она, эта модель, удобна,но только как исходная позиция, когда дешифрован (неточно и не до конца) первичный уровень кодонов иРНК, уровень вещественно-матричных геносигналов, составляющих 1 - 5% от всей массы геномной ДНК. Оставшаяся большая часть ДНК, существующая в понимании большинства генетиков в качестве "мусорной", несет, вероятно, стратегическую информацию о биосистеме в форме потенциальных и действительных волновых сигналов солитонной, голографической и иной образно-знаковой, в том числе и рече-подобной структуры (подробно см. выше главу “Пересмотр модели генетического кода”).
Вероятно, в прямой связи со всеми рассмотренными “аномальными” свойствами генома высших биосистем стоит феномен особого рода, требующий пристального внимания. Это проблема происхождения жизни, и в частности на Земле. Обсуждается она давно. Предположений много. Мы придерживаемся гипотезы панспермии, но не в том варианте, что на Землю были занесены некие споры-родоначальники всех жизненных форм. Вероятно, процесс естественной эволюции абиогенно возникшего “первичного бульона” из органических молекул - предшественников РНК, ДНК, белков и других существенных компонентов биосистем был сочетан с актом введения экзобиологической информации в первые нуклеиновые кислоты, она была артефактом. И эта информация была рече-подобной. “В начале было слово...”. И эти слова были фрактальны, условно начиная с дуплетно-триплетного кода ДНК-РНК, на первых этапах являющегося простейшим языком с четырех буквенной азбукой. Далее произошла трансляция в 20-буквенную азбуку белков и в более высокие языки в духе обсуждавшихся идей. Вообще гипотеза артефакта первичного языка ДНК широко обсуждается, начиная с пионерской работы В. И. Щербака, показавшего искусственность (привнесенность извне) коллективных симметрий генетического кода, вероятность эволюционного происхождения которых близка к нулю. Можно солидаризироваться с такой позицией не только по причине ее красоты и изящного способа доказательств, где в качестве реперных единиц теоретического анализа используются такие параметры, как нуклонные соотношения в аминокислотах и вырожденность генетического кода, но и потому, что она хорошо соответствует нашему мышлению. Однако, введем поправку. Поскольку на самом деле генетический код, то есть код биосинтеза белков, существенно отличается от принятого в начале 60-х г. хх века (см. выше), то и концепция артефакта кода также нуждается в уточнении. Можно предсказать в истинном (фрактальногетеромультиплетном) коде наличие и других знаковых математических образований, фрактально увеличенных по сравнению с теми, что открыл В.И. Щербак.
Развивая эту мысль и ранее выдвинутые нами идеи, скажем, что было бы наивным упрощением считать “языки” и “письменность” (“речь”) ДНК полным аналогом вербальных построений человека. Точнее будет полагать, что функции ДНК основаны прежде всего на ее метаязыке, являющимся грамматикой генома. Здесь чрезвычайно полезен анализ метаязыков А.Соломоником. Математика - тоже метаязык, он же и свод правил построения ее текстов. В отличие от обычной речи, в которой фразу с определенной мыслью можно сконструировать десятками разных способов, в математике ее вербальные (знаковые) ряды генерируются с помощью малого количества жестких правил. И они, правила, позволяют в автоматическом режиме получить предсказательный результат, как в нашем случае с антенной моделью, “предугадывающей” характер резонансных взаимодействий физических полей с информационными биомакромолекулами. На этом примере видно, как в конечном пункте математических метаязыковых (грамматически ориентированных) преобразований получается результат в форме физико-математического образа потенциального поведения важнейших компонентов биосистемы в ее полевом окружении и внутренней наполненности волновыми процессами. Хромосомы, возможно, также оперируют метаязыками для создания “идеальной” (физико-химико-математической) модели биосистемы как практически недостижимого прообраза реального организма. И такая модель будет более информативна по сравнению, например, с голографической моделью, и будет дополнять последнюю.
Если ДНК, хромосомы организмов Земли действительно являются одновременно донорами и акцепторами не только собственных волновых команд, но и неких внешних (возможно, экзобиологических) регуляторных волновых влияний, что было показано нами ранее, то новый искусственный, создаваемый людьми, электромагнитный семиотический канал вхождения в ноосферу и генофонд планеты Земля требует сверхвнимания в отношении уровня разумности и целесообразности наших, по сути неконтролируемых, супергенетических манипуляций. В этом случае мы будем входить в конкуренцию с вероятным экзобиологическим контролем. Полезно ли это и нужно ли? Сейчас ясного ответа нет. Возможно, мы вошли в бифуркационную вилку выбора стратегии эволюции человечества - или идти дальше по техногенному пути, или учиться мудрости у собственного тела, в котором сосредоточена мудрость Творца.
В качестве иллюстрации предложенного нами метода фрактального представления естественных и генетических текстов приведены матрицы плотности для текста на английском языке (руководство по программированию) и “текста” гена казеина (Cazein). Этот метод дает принципиально иную возможность количественного и качественного сравнения естественных и генотекстов. Аналогичный результат можно получить по- иному, и также новым методом, как это показано на графиках гистограмм сходства и различия для фланков и интронов большой группы генов. Таким же путем получена гистограмма сравнения естественных текстов для монографии автора “Волновой геном” и рассказов Ф.Абрамова [Неопубликованные результаты совместных исследований в соавторстве с М.Ю. Масловым (Математический институт РАН)].
Матрица плотности хаотически-игрового представления нуклеотидной последовательности (ген) в алфавите (A,T,G,C), кодирующей первичную структуру казеина (белок молока).
Рис. 11

Матрица плотности хаотически-игрового представления текста на английском языке (руководство по компьютерному программированию). Рассматривалась структура появления в тексте четырёх частей речи. Левый ближний угол соответствует слову “the”, правый ближний - слову “in”, левый дальний - “on”, правый дальний - ”of”.
Рис. 12

Сравнение фланков с интронами
Рис. 13
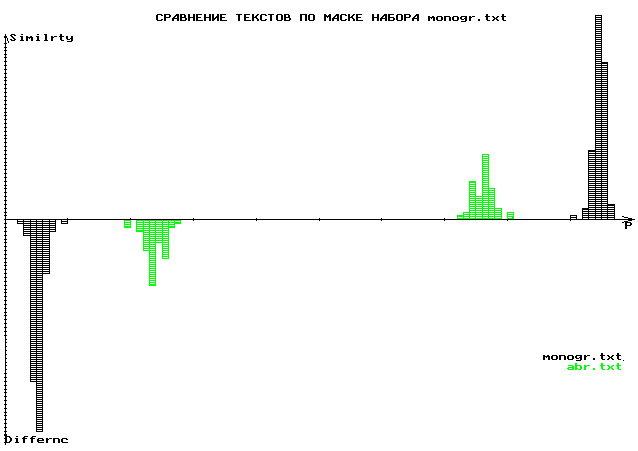
Сравнение монографий:Гаряев П.П. Волновой геном. М.,1994.
и Абрамов Ф. Были небыли. Рассказы. М., 1993.
Рис. 14
В международном компьютерном еженедельнике “Сomputer World” (№ 5 от 3 октября 1995 г.) в рубрике “Подробности” была опубликована подборка статей, посвященная работам по созданию биокомпьютера на главной генетической молекуле - на ДНК. Томас Хоффман в статье “Болотная электроника...” описывает первые робкие попытки использования информационных биомакромолекул - некоторых белков (бактерио-родопсин, родопсин) в качестве субстратов записи-считывания информации как аналогов оптической дисковой памяти. Без сомнений, это интересное оригинальное направление, однако в данном случае ничего принципиально нового предложено не было, поскольку неважно откуда взято вещество-субстрат записи-считывания информации, на котором получают спектральные выжигания типа двоичного кода или с помощью лазеров записывают трехмерные изображения предметов в форме голограмм. Такое вещество может иметь абиогенное происхождение или, как в случае с родопсинами, извлекаться из биомембран солончаковых бактерий. В связи с этим, учитывая наши исследования, было бы логичным рассматривать молекулы ДНК как неразрывное единство Вещества и Поля также и в аспекте их участия как основной рабочей фигуры в искусственных биокомпьютерах. Это было бы полезно в развитии вычислительной техники и может привести к полной смене ее элементной базы в ряду: аналоговый-цифровой-“образный” или смысловой компьютер на ДНК.
Весной 1995 г. Леонард М. Адлеман, профессор вычислительных наук из Университета Южной Калифорнии, описал в журнале “Science” алгоритм использования ДНК для решения одной из версий “задачи коммивояжера”. Потребовалась всего неделя для получения ответа, в то время как традиционным компьютерам понадобилось бы несколько лет. При этом было использовано фундаментальное явление, свойственное молекулам ДНК - способность к так называемым комплементарным взаимоузнаваниям. Это явление заключается в том, что любые фрагменты каждой из двух цепочек ДНК находят в растворе (или в составе хромосом живой клетки) только собственные, в некотором смысле зеркальные, половинки и образуют нормальную двойную спираль. Успешность и быстрота автоматических поисков половинками ДНК друг друга как акта самоорганизации (самосборки) и обеспечили высокую скорость перебора вариантов в пределах “задачи коммивояжера”. Причины быстрых и точных взаимоузнаваний половинок ДНК до недавнего времени были неизвестны. А это необычайно важно для реального создания ДНК-компьютера, и об этом речь пойдет ниже.
Путь, который выбрал Адлеман, используя ДНК, не то чтобы неверен, скорее, он похож на попытки понять, как, например, происходит процесс мышления у Иванова, Петрова или Сидорова на основе нашего знания о том, что они любят вкусно поесть. Правильное и эффективное использование ДНК, как основного информационного элемента будущего биокомпьютера, немыслимо без понимания истинных функций генетических молекул в биосистемах. Возвращаясь к предыдущим главам, хромосомный аппарат, как система записывающая, сохраняющая, изменяющая и транслирующая информацию, может рассматриваться одновременно на уровнях вещества и достаточно хорошо изученных физических полей, которыми, как носителями генетической и общерегуляторной информации, оперирует континуум генетических молекул (ДНК,РНК). Континуум этот является основным компонентом совокупности хромосом, являющейся, по сути, биокомпьютером. Уровни вещества и поля, на которых хромосомный биокомпьютер функционирует, неразрывны и функционально дополняют друг друга. Здесь реализуются неизвестные ранее виды памяти (солитонная, голографическая, фантомная) и при этом молекулы ДНК могут работать как биолазеры и одновременно как среда записи лазерного сигнала. Кроме того, мы обнаружили, что ДНК способна излучать широкополосное сверхслабое электромагнитное поле, которое нам удалось усилить в тысячи раз. Впрочем, ДНК в этом плане является частным случаем, поскольку зафиксированное нами явление свойственно, вероятно, всем веществам [42], но хромосомы используют этот феномен, наверное, в высшей степени эффективно как один из волновых каналов информационных и (или) энергетических коммуникаций. Молекулы ДНК, как континуум любой биосистемы, способны к формированию прообразов биоструктур и организма в целом как “волновых копий” или “матриц” и сравнению построенного организма с ними как с реперами. В этом плане механизм быстрого и точного взаимоузнавания цепочек (половинок) ДНК, механизм, которым воспользовался Адлеман для решения “задачи коммивояжера”, - лишь один из способов самоорганизации биосистем. Взаимоузнавание, в частности, происходит потому, что в молекулах ДНК зарождаются особые сверхустойчивые акустико-электромагнитные волны (так называемые солитоны), некоторые разновидности которых можно трактовать в рамках открытого в 1949г. “явления возврата Ферми-Паста-Улама” (ФПУ). Такие солитоны ДНК обладают двумя связанными типами памяти - собственно памятью, свойственной явлению ФПУ-возврата, т.е. способностью помнить начальные моды возбуждений и периодически к ним “возвращаться”. Другая память ДНК-континуума в биосистеме - квази-голографическая или фрактальная. Она связана с фундаментальным свойством биосистем - восстанавливать целое из своей части. Это свойство фудаментально и хорошо известно (черенкование растений, регенерация хвоста у ящериц, регенерация целого организма из яицеклетки). Высшая форма такой памяти - ассоциативная память коры головного мозга, т. е. нейронов. Бесперспективно рассуждать о ДНК-компьютере, даже решив с помощью молекул ДНК “задачу коммивояжера”, если не учитывать новую логику в понимании знаковых, кодирующих биофункций ДНК. Другая сторона дела состоит в соотнесении этой логики с многочисленными исследованиями по нейрокомпьтерам и попытками разобраться в “компьютерной” работе мозга без понимания кодирующих функций нервного импульса.
Принято считать, что нервный импульс прост, что он является всего лишь волной деполяризации, и поэтому шифровка информации здесь происходит только по частотному механизму. Расчеты показывают, однако, что частотные модуляции недостаточны для кодирования. А.А.Березиным проведено крупное исследование, из которого следует, что нервные импульсы - это все те же солитоны в рамках явления ФПУ-возврата, при этом, самое главное, такие солитоны отображают в своей структуре РНК-”тексты”, а следовательно, и ДНК В развитии этой идеи нами выдвинуто предположение о знаковой смысловой связи солитонов на хромосомном уровне и солитонов нервных импульсов. Это свидетельствует в пользу связи сознания и его отображения - слова с основной информационной фигурой биосистем - молекулами ДНК с их новыми, неизвестными ранее, типами памяти. Солитоны ДНК способны после “прочитывания ими генотекстов” покидать пределы ДНК в форме особых электромагнитных полей с тем, чтобы узнавать нужные участки других молекул ДНК и формировать целостные новые “тексты”, необходимые в данный момент для выполнения биофункций, в том числе и путем передачи информации в нервные импульсы. Что касается термина “тексты ДНК”, который раньше употреблялся для удобства и был взят взаймы у лингвистов для метафорического употребления, то оказывается, эта текстовая структура ДНК действительно сродни человеческой речи. Наши математико-лингвистические исследования [14,25, см. также предыдущие главы] показали, что такой ключевой параметр, как фрактальность, един для ДНК и человеческой речи. Это коррелирует с ранними работами в этой области
Используя эти теоретические разработки и собственные данные по физико-химии ДНК, нам удалось экспериментально доказать возможность свертки генетической информации в форме солитонных волновых пакетов, описываемых физико-математическим формализмом явления возврата Ферми-Паста-Улама (ФПУ). Такие волновые пакеты с искусственно введенной в них биоинформацией, продуцируемые соответствующими ФПУ-радиоэлектронными устройствами, способны входить в резонансный информационный контакт с генетическим аппаратом животных, растений и, вероятно, человека с последующим резким и направляемым изменением их обмена веществ. Оказалось, что вещество наследственности - ДНК - является генератором ФПУ-солитонных акустических и электромагнитных полей. Именно поэтому ФПУ-генераторы способны вводить информацию в хромосомы по электромагнитным резонансным механизмам. Эффективность генераторов на порядки возрастает, если практически использовать феномен математической общности фрактальной структуры ДНК-”текстов” и человеческой речи. Грамматика генетических текстов является, вероятно, частным случаем универсальных грамматик всех языков людей. Поэтому и реализуются физико-смысловые резонансы солитонных структур ДНК и искусственных ФПУ-солитонных полей, как аналогов естественных ФПУ-хромосомных полей.
Вводя определенные кодовые вербальные команды через генератор ФПУ в генетический аппарат радиационно поврежденных семян растений, удалось достоверно уменьшить число хромосомных аберраций, т.е фактически блокировать поражающее действие рентгеновского облучения. Более того, оказалось, что возможна превентивная защита генома растений от жесткого рентгеновского излучения с помощью адекватных волновых команд. Контрольные эксперименты с хаотическими вербальными построениями (командами), введенными через ФПУ-устройства в геном биосистем, показали, что такие команды никак не влияют на целостность хромосом.
Эти эффекты предсказаны и проверены на основании использования математических компьютерных моделей, имитирующих “чтение” солитонами на ДНК генотекстов и ретрансляцию этих текстов в другие клетки и ткани.
Другие наши физико-математические модели и эксперименты обосновывают так называемый “антенный эффект” при возбуждении электромагнитными полями выделенных коллективных мод макромолекул ДНК. Это прямо связано с экспериментами по так называемому двухфотонной накачке геноструктур с последующим лазерным излучением ДНК [18] и также согласуется с нашими результатами по взаимодействию гелей ДНК с импульсным излучением инфракрасного лазера [25].
Вновь вернемся к компьютеру на ДНК. Ясно, что при его разработке необходимо использовать не только и не столько результаты эксперимента Адлемана. Чтобы реализовать свои возможности, ДНК должна находиться в привычной среде - в водном растворе или в жидкокристаллическом состоянии. Но это лишь начало. Другие возможности ДНК или хромосом могут быть выявлены в условиях, приближенных к тем, которые имеются в живой клетке. В пределе компьютер на ДНК - это и есть живая клетка, то есть надо создать искусственный организм, а это произойдет не скоро. Сейчас мы можем делать только какие-то приближения к состоянию ДНК в клетке, но и это немало. Можно перечислить то, что реально выполнимо уже сейчас. Прежде всего необходимо начать практическое использование новых типов памяти геноструктур и для этого пытаться конструировать ячейки памяти, работающие на явлении ФПУ-резонансов и (или) на способности записывать голограммы. Такая память будет на многие порядки по объему и быстродействию превосходить память существующих магнитных, оптических дисков и голографических систем. Вторая принципиальная возможность связана с этими типами памяти, но многократно усиливается способностью хромосом быть лазеро-активной средой. Препараты хромосом выступают в таком варианте и как ячейка памяти, и как лазеры, считывающие собственную (а также наведенную) голографическую и ФПУ-память. И, наконец, последняя из достижимых в настоящее время - использование квази-речевых характеристик ДНК. Можно создавать такие ДНК-лазеры, которые будут высвечивать и “озвучивать” как естественные генотексты, так и искусственные (синтезированные) знаковые последовательности полинуклеотидов, имитирующие естественные генопрограммы. Однако это весьма опасный путь и необходима система запретов на искусственные волновые гены. Такой способ работы с ДНК-компьютерами означает вхождение в новые семиотические ареалы генома человека, вообще всей биосферы, ареалы, которые Природа (или Бог) использовала для создания человека. Мысль вполне реалистичная, если учесть теоретические работы по коллективной симметрии генетического кода, проводимые школой Эйгена в Институте Макса Планка. Ее исследования показывают, что ключевая часть информации, записанная и записываемая как квазиречь в хромосомах всех организмов нашей планеты, носит искусственный характер. Наши данные о том, что хромосомный континуум и ДНК любой биосистемы является неким подобием антенны, открытой во вне для приема дополнительной (возможно, экзобиологической) информации, подтверждают сказанное. Можно думать, что геном организмов Земли, по крайней мере частично, является полигоном для смысловых экзобиологических влияний, и в этом плане существенно, что мы нашли первичные подходы к вхождению в этот семиотико-семантический ареал.
Основываясь на сказанном, можно предсказать, что открываются следующие перспективы знаковых манипуляций с геноструктурами как основным субстратом биокомпьютеров:
1. Создание искусственной памяти на генетических молекулах, обладающей поистине фантастическим объемом и быстродействием.
2. Создание биокомпьютера на ДНК, основанного на совершенно новых принципах и сравнимого по способам обработки информации и функциональным возможностям с человеческим мозгом.
3. Осуществление дистантного управления ключевыми информационными процессами в биосистемах через искусственные биокомпьютеры (лечение рака, СПИДа, генетических уродств, управление социогенетическими процессами и, в конечном итоге, изменение времени жизни).
4. Активно защищаться от деструктивных волновых влияний через обнаруженный информационно-энергетический канал.
5. Устанавливать экзобиологические контакты.
Подводя итог, спросим : что остается от логики постановочных экспериментов с ДНК, которую предлагают Адлеман и другие исследователи в области молекулярной электроники информационных биомакромолекул? Эта логика уязвима, поскольку основана на упрощенных представлениях о работе хромосом как чисто вещественного субстрата. Волновые функции геноструктур не берутся в расчет. Это тупик, который оборачивается все более нарастающим огромным финансированием по гено-биотехнологиям, по нейрокомпьютерам со все меньшим практическим выходом. Те же пороки ожидают и молекулярную электронику в ее попытках использовать одномерное мышление относительно ДНК при создании биокомпьютера.
Такой компьютер должен имитировать функции генома в части оперирования волновой информацией - то есть создавать образы, в том числе и квази-речевые, распознавать их, манипулировать ими как командными. Такие знаковые структуры будут обладать огромной биологической активностью. Даже сейчас лазер на ДНК, “заряженный” определенными текстами, например, геном продолжительности жизни, вероятно, мог бы продлить ее у человека на 300 - 400 и более лет. Необходимо перераспределение финансирования в генетике, эмбриологии и генной инженерии, а также в молекулярной электронике. Это позволит сделать прорыв в создании компьютеров с квази-генетической памятью, объем которой превосходит все мыслимые пределы, и способных управлять суперсложными процессами, реально сравнимыми с метаболизмом и мышлением. Такие биокомпьютеры будут способны контролировать и поддерживать нормальную жизнедеятельность человека во временных масштабах, приближающихся к бессмертию.
Нами обнаружено явление перехода красного когерентного света в радиоволны при взаимодействии лазерного пучка с веществами. Рассмотрим, как формируются на порядки усиленные радиоизлучения геноструктур in vitro, формируемые за пределами лазерного резонатора, основной частью которых могут являться электроакустические поля, обнаруженные нами ранее, в частности, как волновые структуры ДНК, рибосом, коллагена и хромосом [25].
Для получения стоячей и бегущих волн на лазерном пучке, зондирующем геноструктуры, в наших экспериментах был использован специально изготовленный He-Ne лазер с мощностью излучения 2 mВт, длиной волны 632,8 нм, одночастотный со стабильным резонатором, управляемым посредством термостатирующего элемента. Схема эксперимента приводится на рис 15.
Это схема так называемого трехзеркального лазерного интерферометра имеет особенность в том, что внешнее зеркало резонатора является полупрозрачным или полностью прозрачным, а также может быть преобразовано в сложные композиции типа “сэндвич”, между слоями которого вводятся изучаемые препараты. В этой схеме имеются два информа-
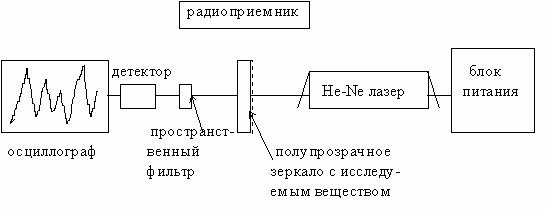
Рис.15
ционных канала - один традиционный оптический, а другой по эфиру - радиоволновый. Рассмотрим вначале работу оптического канала для того, чтобы понять работу второго канала. Внешнее зеркало резонатора юстируется таким образом, чтобы отраженная часть оптического луча от этого зеркала точно попадала бы в резонатор. При этом прямой из лазера и отраженный от зеркала лучи интерферометра складываются и результирующая волна от сложения сигналов, прошедшая через полупрозрачное зеркало, анализируется пространственным фильтром и попадает на кристалл фотодетектора. Пространственный фильтр, представляющий собой точечную диафрагму, жестко соединен в едином моноблоке с корпусом фотодетектора, электрический сигнал от которого наблюдается на экране осциллографа. В зависимости от того, в какой спекл попадает точечный участок фильтра, наблюдаются те или иные осциллографические сигналы, которые появляются за счет резонансного усиления радиоизлучений, формируемых за пределами лазерного резонатора.
Регистрацию радиоволн проводили
“радиопрослушиванием” близлежащего
пространства (в радиусе до 4-х метров) вокруг
препаратов специальным образом приготовленных
хромосом (сперматозоиды мыши) и ДНК из селезенки
быка. Концентрированные препараты (около 1 мг/мл в
дистиллированной воде, а также в определенных
сочетаниях с этанолом, водой, триптофаном и
гуанозин-трифосфатом) наносили тонким слоем на
грань полупрозрачного зеркала (предметное
стекло), накрывали вторым предметным стеклом,
выдерживали 3 - 4 суток до высыхания при ![]() , и направляли отраженный
(промодулированный исследуемым препаратом)
пучок света обратно в лазерный резонатор. В
другой модификации использовали тонкую,
отражающую свет, пленку ДНК без нанесения ее на
полупрозрачное зеркало. Схема эксперимента с
воздействием ультразвука на ДНК на рис.16.
, и направляли отраженный
(промодулированный исследуемым препаратом)
пучок света обратно в лазерный резонатор. В
другой модификации использовали тонкую,
отражающую свет, пленку ДНК без нанесения ее на
полупрозрачное зеркало. Схема эксперимента с
воздействием ультразвука на ДНК на рис.16.
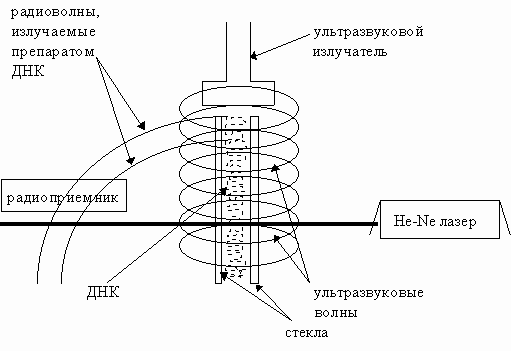
Рис.16
При всех способах подготовки ДНК
регистрировали отчетливые радиосигналы,
различающиеся по характеру в зависимости от типа
исследуемых образцов или их сочетаний.
Зондирование ДНК и хромосом и их комплексов с
перечисленными выше веществами сопровождалось
особыми радиосигналами (радиозвуком), резко
отличным от такового абиогенных препаратов.
Характерным, и полностью совпадающим с данными
[25], было наличие чередования четких одиночных
радио звуковых периодических (или почти
периодических) сигналов, чередующихся со
стохастическими, аналогично наблюдению в [25]
относительно характера воздействия
инфракрасного лазера ![]() на ДНК. Представляется, что это еще одна
демонстрация высокой самоорганизации
(солитонообразования) ДНК в рамках явления
возврата Ферми-Пасты-Улама, самоорганизации,
свойственной генетическим структурам как одно
из фундаментальных проявлений памяти
наследственности. Иными словами, в настоящем
исследовании мы в какой-то степени подтвердили
обнаруженную в [25] “солитонную память” ДНК на
определенные моды возбуждения препарата,
например, на механические вибрации. Характерно,
что специфические модуляции радио звука при этом
полностью соответствовали изменению во времени
двумерных спекл-картин рассеянного препаратами
ДНК света. Этот эффект полностью прекращался,
если на препараты воздействовали ультразвуком (25
кГц, мощность 6,6
на ДНК. Представляется, что это еще одна
демонстрация высокой самоорганизации
(солитонообразования) ДНК в рамках явления
возврата Ферми-Пасты-Улама, самоорганизации,
свойственной генетическим структурам как одно
из фундаментальных проявлений памяти
наследственности. Иными словами, в настоящем
исследовании мы в какой-то степени подтвердили
обнаруженную в [25] “солитонную память” ДНК на
определенные моды возбуждения препарата,
например, на механические вибрации. Характерно,
что специфические модуляции радио звука при этом
полностью соответствовали изменению во времени
двумерных спекл-картин рассеянного препаратами
ДНК света. Этот эффект полностью прекращался,
если на препараты воздействовали ультразвуком (25
кГц, мощность 6,6 ![]() ) в
течение 10-15 секунд на расстоянии 1-2 сантиметра.
После этого радиозвук становился монотонным и
практически не отличался от фонового.
) в
течение 10-15 секунд на расстоянии 1-2 сантиметра.
После этого радиозвук становился монотонным и
практически не отличался от фонового.
Можно высказать рабочую гипотезу о том, какие события происходят при взаимодействии ультразвука и генетических структур:
а) происходят разрывы молекул ДНК;
б) некоторые моды (или обертона) “записываются” на уровне акустики ДНК в рамках солитонного явления возврата Ферми-Паста-Улама и периодически возвращаются, “проигрываются” геноструктурами, подавляя (зашумляя) генознаковую ( в том числе и онко-генознаковую) акустику ДНК;
в) происходят оба перечисленных процесса.
В живом организме при хирургических операциях на раковых опухолях ультразвуковыми скальпелями, разработанных В.И.Лощиловым, это приводит к “стиранию” извращенной генетической информации, даваемой онкогенами, и тем самым к прерыванию метастазов.
Это служит основой для разработки принципиально новых методологий “волновой хирургии” онкологических больных, а если шире, то и “волновой медицины”.
Попытаемся дать первичную физическую
модель феномена, когда лазерный свет (при
взаимодействии с исследуемыми веществами)
генерирует радиоволны. В общем случае всякую
излучающую систему можно представить в виде
потенциального гармонического осциллятора,
находящегося в первоначальном состоянии
устойчивого равновесия. В нашем случае, когда
внешнее зеркало резонатора неподвижно, система
находится в состоянии устойчивого равновесия и
не излучает радиоволны. При этом (![]() ), для которого потенциал
), для которого потенциал ![]() минимален. При отклонении
зеркала из устойчивого равновесия на некоторое
расстояние
минимален. При отклонении
зеркала из устойчивого равновесия на некоторое
расстояние ![]() потенциал в
этой точке может быть разложен в ряд по степеням
малых величин
потенциал в
этой точке может быть разложен в ряд по степеням
малых величин ![]() , а так
как в этой точке частная производная
, а так
как в этой точке частная производная ![]() = 0, то будем иметь значения
для потенциала гармонического осциллятора
= 0, то будем иметь значения
для потенциала гармонического осциллятора
![]()
![]() .
.
Запишем волновое уравнение для гармонического осциллятора в виде:
![]() , (1)
, (1)
где ![]() -
постоянная планка,
-
постоянная планка,
![]() -
квазиупругая постоянная,
-
квазиупругая постоянная,
![]() , где
, где ![]() - угловая частота
колебаний, а
- угловая частота
колебаний, а
![]() -потенциал.
-потенциал.
Если 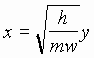 ,
, ![]() , то уравнение (1) примет
вид:
, то уравнение (1) примет
вид:
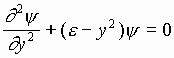 .
.
Известно, что энергия гармонического
осциллятора всегда положительна, и для среднего
значения энергии ![]() имеем:
имеем:
![]() ,
,
где собственные значения ![]() будут:
будут:
![]() ,
,
и они равны собственным значениям
гармонического осциллятора с точностью для
полуцелого квантования энергии. В нашем случае
имеет место суперпозиция двух (![]() и
и ![]() )
монохроматических гармонических волн от
точечных осцилляторов, одним из которых является
лазер, а другим внешнее зеркало резонатора. Для
упрощения выкладок построения
физико-математической модели предположим, что
поляризация этих волн одинакова и совпадает по
ориентации магнитных
)
монохроматических гармонических волн от
точечных осцилляторов, одним из которых является
лазер, а другим внешнее зеркало резонатора. Для
упрощения выкладок построения
физико-математической модели предположим, что
поляризация этих волн одинакова и совпадает по
ориентации магнитных ![]() и
электрических
и
электрических ![]() векторов
Умова-Пойтинга. Результирующее колебание в такой
суперпозиции (
векторов
Умова-Пойтинга. Результирующее колебание в такой
суперпозиции (![]() и
и ![]() ) волн дадут:
) волн дадут:
![]() ,
,
где ![]() и
и ![]() - амплитуды волн;
- амплитуды волн; ![]() и
и ![]() - фазы;
- фазы; ![]() -
угловая частота;
-
угловая частота; ![]() -
время.
-
время.
Для волн, распространяющихся в одном направлении, результирующее колебание можно записать как
![]() ,
,
где
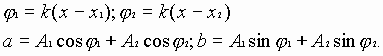 (2)
(2)
Если ![]() , то
, то ![]() .
.
Из уравнения (2), получим новую гармоническую волну, амплитуда и фаза которой будет иметь вид:
![]() ,
,
![]() =
=![]() . (3)
. (3)
Угол (![]() ) в
выражении для амплитуды результирующей волны
равен
) в
выражении для амплитуды результирующей волны
равен
(![]() )
)![]() .
.
Таким образом, произведя сложение волн
посредством третьего зеркала резонатора,
которое луч возвращает точно в резонатор лазера,
получили новую гармоническую волну. Из
последнего выражения видно, что амплитуда не
зависит от положения точки, в которой мы
исследуем результирующее колебание. Все точки
колеблются с одинаковой амплитудой, зависящей
лишь от разности расстояний между точками, в
которых фазы суммируемых волн равны нулю.
Амплитуда будет максимальной, когда разность фаз
будет равна ![]()
![]() , где
, где ![]() - целое число. При этом расстояние между
внешним зеркалом и резонатором лазера составит
- целое число. При этом расстояние между
внешним зеркалом и резонатором лазера составит ![]() , а амплитуда равна
, а амплитуда равна ![]() . В этом случае волны
складываются в фазе. Минимальной амплитуда будет
при условии, если
. В этом случае волны
складываются в фазе. Минимальной амплитуда будет
при условии, если ![]()
![]() , а разность путей двух
волн
, а разность путей двух
волн ![]() . В этом случае
амплитуда равна
. В этом случае
амплитуда равна ![]() ,
что соответствует сложению противофазных волн.
Если амплитуды общих волн будут равны, т.е.
,
что соответствует сложению противофазных волн.
Если амплитуды общих волн будут равны, т.е. ![]() , то наименьшая амплитуда
будет равна нулю, а максимальная - удвоенной
амплитуде суммируемых волн. При равенстве
амплитуд фаза результирующего колебания будет
равна
, то наименьшая амплитуда
будет равна нулю, а максимальная - удвоенной
амплитуде суммируемых волн. При равенстве
амплитуд фаза результирующего колебания будет
равна ![]() и не будет
зависеть от положения точки наблюдения. Во всех
точках пространства, где происходит
интерференция, фаза колебания одинакова, что
соответствует стоячим волнам.
и не будет
зависеть от положения точки наблюдения. Во всех
точках пространства, где происходит
интерференция, фаза колебания одинакова, что
соответствует стоячим волнам.
Амплитуда этих волн из (3) ![]() . В случае встречных волн с
различными амплитудами
. В случае встречных волн с
различными амплитудами ![]() .
При наложении таких волн образуется стоячая
волна с амплитудой
.
При наложении таких волн образуется стоячая
волна с амплитудой ![]() , а
также бегущие волны с амплитудой
, а
также бегущие волны с амплитудой ![]() , которые распространяются в
направлении волны с большей амплитудой
, которые распространяются в
направлении волны с большей амплитудой ![]() . Эти бегущие волны при
записи создают фон, над уровнем которого
наблюдается модуляция, обусловленная “дышащей”
стоячей волной типа трубчато-цилиндрического
солитона-бризера. При неподвижном зеркале
внешнего резонатора в нашем случае образуется
трубчато-цилиндрический квантовый
солитон-бризер, стоящий на месте, но “дышащий”.
. Эти бегущие волны при
записи создают фон, над уровнем которого
наблюдается модуляция, обусловленная “дышащей”
стоячей волной типа трубчато-цилиндрического
солитона-бризера. При неподвижном зеркале
внешнего резонатора в нашем случае образуется
трубчато-цилиндрический квантовый
солитон-бризер, стоящий на месте, но “дышащий”.
В нелинейной физике такого рода
солитоны названы бризерами именно потому, что
они, оставаясь на месте или смещаясь вблизи
неоднородностей (например, зеркала), “дышат” (от
английского breath). Именно это низкочастотное
“оптическое солитонное дыхание”, как нам
представляется, и генерирует радиоволны ![]() и
и ![]() . “Солитонное акустико-электромагнитное
дыхание” свойственно и молекулам ДНК, рибосомам
и коллагену в рамках явления возврата
Ферми-Паста-Улама. [25]. Колебания
жидкокристаллической решетки молекул ДНК
приводят к возникновению поперечных волн
интенсивности, проявляющихся в изменении
диаметра трубчато-цилиндрического квантового
солитона-бризера, который может двигаться,
порождая широкий спектр радиоволн. Диаметр этого
солитона может уменьшаться или увеличиваться
при прямом или обратном движении внешнего
зеркала резонатора. Систему подвижных колец
(проекций трубок), в той или иной степени
искаженных (промо-дулированных) исследуемым
препаратом, можно наблюдать на экране,
установленном между внешним зеркалом резонатора
и пространственным фильтром фотодетектора.
Движение такого солитона-бризера может привести
к усилению тех бегущих волн интенсивности, о
которых речь шла выше. Таким образом, происходит
взаимная энергетическая подпитка бегущих волн
солитоном-бризером и наоборот. Можно полагать,
что подкачка расходуемой энергии
солитона-бризера происходит из фоновой
стохастической энергии kT за счет
стохастического резонанса через фильтр
согласования между кратными гармониками макро- и
микросистем, где макросистема есть
солитон-бризер, а микросистемой является
отдельный квантовый осциллятор, например,
молекула ДНК.
. “Солитонное акустико-электромагнитное
дыхание” свойственно и молекулам ДНК, рибосомам
и коллагену в рамках явления возврата
Ферми-Паста-Улама. [25]. Колебания
жидкокристаллической решетки молекул ДНК
приводят к возникновению поперечных волн
интенсивности, проявляющихся в изменении
диаметра трубчато-цилиндрического квантового
солитона-бризера, который может двигаться,
порождая широкий спектр радиоволн. Диаметр этого
солитона может уменьшаться или увеличиваться
при прямом или обратном движении внешнего
зеркала резонатора. Систему подвижных колец
(проекций трубок), в той или иной степени
искаженных (промо-дулированных) исследуемым
препаратом, можно наблюдать на экране,
установленном между внешним зеркалом резонатора
и пространственным фильтром фотодетектора.
Движение такого солитона-бризера может привести
к усилению тех бегущих волн интенсивности, о
которых речь шла выше. Таким образом, происходит
взаимная энергетическая подпитка бегущих волн
солитоном-бризером и наоборот. Можно полагать,
что подкачка расходуемой энергии
солитона-бризера происходит из фоновой
стохастической энергии kT за счет
стохастического резонанса через фильтр
согласования между кратными гармониками макро- и
микросистем, где макросистема есть
солитон-бризер, а микросистемой является
отдельный квантовый осциллятор, например,
молекула ДНК.
Первоначальная идея использования энергии kT для подпитки солитонов обсуждена в работе Пиппарда и в наших последующих исследованиях относительно создания лазеров на информационных биомакромолекулах, в частности, лазеров с использованием солитонных волн [2,3,16,34,35]. Фильтр согласования мы понимаем как инструмент реализации общего принципа гармонического сочетания, то есть непротиворечивого согласованного взаимодействия. Вычислим диапазон спектра частот генерации такого опто-радиоволнового “дыхания” в наших экспериментах за счет движения внешнего зеркала резонатора. Спектр частот резонатора определяется временем прохождения излучением всего тракта от торца дальнего зеркала лазера до внешнего зеркала резона-
тора по известному соотношению:
 ,
,
где ![]() - целое
число;
- целое
число;
![]() - длина и
показатель преломления среды
- длина и
показатель преломления среды ![]() -го участка резонатора;
-го участка резонатора;
![]() - длина
резонатора;
- длина
резонатора;
![]() -скорость
света в среде.
-скорость
света в среде.
В резонаторе, как правило, существуют
несколько типов бегущих волн, для которых число ![]() различается на единицу и
зависит от количества мод. Такой режим является
многомодовым. Разность частот между соседними
модами составляет
различается на единицу и
зависит от количества мод. Такой режим является
многомодовым. Разность частот между соседними
модами составляет
В резонаторе резко сужается ширина спектра излучения. Известно, что добротность резонатора определяется соотношением
![]() ,
,
где ![]() -
энергия, запасенная в резонаторе;
-
энергия, запасенная в резонаторе;
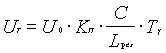 - это
энергия, теряемая в резонаторе за цикл времени
одного прохода резонатора
- это
энергия, теряемая в резонаторе за цикл времени
одного прохода резонатора ![]() ,
,
где ![]() -коэффициент
суммарных потерь;
-коэффициент
суммарных потерь;
![]() -потери
энергии за один проход в резонаторе;
-потери
энергии за один проход в резонаторе;
![]() - время
прохода резонатора.
- время
прохода резонатора.
Подставляя значение ![]() в
в ![]() и
учитывая, что
и
учитывая, что ![]() ,
получаем:
,
получаем:
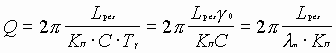 ,
,
так как ![]() , то
, то ![]() ; перепишем
; перепишем
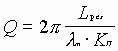 ;
;
Исходя из величины добротности резонатора, находим ширину спектра резонатора:
![]() , (4)
, (4)
который является диапазоном спектра излучения радиоволн.
Из работы известна связь между шириной спектра резонатора и шириной линии излучения:
![]() , (5)
, (5)
где ![]() -
мощность излучения на данной моде;
-
мощность излучения на данной моде;
![]() - энергия
кванта.
- энергия
кванта.
Простой расчет из формулы (4) показывает, что при
![]() =10%;
=10%; ![]() = 0,54м;
= 0,54м;![]() = 632,8нм;
= 632,8нм; ![]() 1мВт
1мВт
![]() и из формулы
(5)
и из формулы
(5) ![]() = 0,007Гц.
= 0,007Гц.
Это трансформированные из лазерного света радиоволны в мегагерцовой области, что и было зарегистрировано в наших экспериментах.
Детальное развитие нашей квантово-электродинамической модели порождения радиоволн из света связано с теорией поля Максвелла и идеей возникновения элементарных частиц, которые обладают ротационной осцилляцией электромагнитных векторов Умова-Пойтинга. Эта модель в дальнейшем позволит углубить понимание механизма перехода от пространства к веществу и наоборот и далее к пересмотру понятия времени. Практическое приложение данной теории позволит объяснить такие “аномальные” и давно известные эффекты, как сверхустойчивость термофильных бактерий к температурам, существенно превышающим области термодинамической устойчивости информационных биополимеров - ДНК и белков, а также “аномально” высокую сопротивляемость некоторых биосистем к огромным уровням радиоактивности. В свою очередь, это может вывести на понимание принципов старения и сопротивления старению организмов через идею согласующих фильтров биосистем и окружающей среды, фильтров, фрактально повторяющих себя на микро- и макромасштабах (от элементарных частицы до бактерий и человека).
Имеется косвенное теоретическое
подтверждение нашей модели генерации радиоволн
из солитонных возбуждений. В работе Тужински и
соавторов показана связь, взаимодополняемость
двух казалось бы независимых теорий, в которых
рассматриваются две физические модели,
объясняющие необычное поведение биологических
систем. Модели предложены Гербертом Фрелихом и
Александром Давыдовым. Так называемые
Давыдовские солитоны, описывающие возбуждение,
делокализацию и движение электронов вдоль
пептидных цепей белковых молекул в форме
уединенных волн (солитонов), дополняют известную
модель Фрелиха, развитую в нашей работе [16] о
возможности высокополяризованного
(когерентного, лазероподобного) состояния
колеблющихся диполей информационных
биомакромолекул, диполей, возникающих при
Бозе-конденсации фононов электромагнитных волн
белков (![]() ), ДНК (
), ДНК (![]() ), мембран (
), мембран (![]() ). В работе [см. сноску 2 на стр. 85]
Давыдовский гамильтониан трансформирован в
нормальные координаты, Фрелиховский
гамильтониан канонически трансформирован в
эквивалентную форму в рамках аппроксимации
Хартри-Фока. Авторы полагают, что модель
гамильтониана способна связать обе теории,
которые математически эквивалентны.
). В работе [см. сноску 2 на стр. 85]
Давыдовский гамильтониан трансформирован в
нормальные координаты, Фрелиховский
гамильтониан канонически трансформирован в
эквивалентную форму в рамках аппроксимации
Хартри-Фока. Авторы полагают, что модель
гамильтониана способна связать обе теории,
которые математически эквивалентны.
Кроме того, обе модели дополняют друг друга физически. Бозе-конденсация вибрационных мод биополимеров соответствует распространению солитона волны поляризации. И наоборот, солитонный транспорт граничной энергии вдоль пептидной цепи сопровождается бозе-конденсацией решеточных вибраций биоструктур.
Отсюда следует, что солитон порождает электромагнитное поле, а это, возможно, тот самый эффект, который мы наблюдаем в экспериментах, когда осциллирующий оптический солитон-бризер, отображающий солитонные возбуждения ДНК, генерирует оптико-резонансно усиленные радиоволны.
Еще одна мысль, привлекающая внимание: конверсия эндогенных когерентных фотонов, генерируемых хромосомами, в радиоволны в биосистеме может происходить по трехзеркальному принципу на многочисленных отражающих поверхностях мембран, аналогично нашим модельным опытам. Именно этим может объясняться зеркальный блеск органов и тканей живых организмов. Это же дает надежду, что мы сумеем in vitro-in vivo манипулировать световыми лазерными потоками, которые проводятся сложнейшей сетью световодов живой клетки и которые преобразуются на биомембранах в знаковые радиоволны. Такие процессы могут использоваться как основа для создания принципиально иных компьютеров, точнее биокомпьютеров.
1. Проведено математическое (компьютерное) моделирование солитонных возбуждений в молекуле ДНК, показавшее способность уединенных волн (на уровне крутильных колебаний нуклеотидов) реагировать на первичную структуру ДНК и тем самым служить потенциальной основой для истантной трансляции текстовой и иной образной управляющей информации генома.
2. Экспериментально зафиксированы солитоноподобные процессы в гелях ДНК in vitro как реализация явления возврата Ферми-Паста-Улама (ФПУ). Тем самым продемонстрирована принципиальная возможность функционирования генетического аппарата биосистем с использованием ФПУ-волнового типа памяти генома и, соответственно, неизвестного ранее способа передачи геноуправляющих сигналов in vivo полевым путем.
3. Экспериментально доказана возможность существования эпигенетической информации, конвертированной в форму широкополосного электромагнитного поля, в котором реализуется пространственно-временной возврат ФПУ. Доказано также, что такая информация может искусственным способом с помощью генератора ФПУ передаваться от биосистемы-донора к биосистеме-акцептору, что соответствует, вероятно, процессам волнового “метаболизма” эпигеносигналов in vivo. Такой способ является, вероятно, примитивной имитацией главного информационного канала, связанного с Творцом.
4. Найдена компьютерная модель фрактального представления, которая показала, что человеческая речь (тексты) и последовательности нуклеотидов (тексты ДНК) обладают близкой математической структурой.
5. Управление развитием высших биосистем происходит с использованием материально-волновых солитонно-голографических матриц генома, а также семиотических единиц, сходных с человеческой речью и, вероятно, с речью Творца.
6. Разработаны технические устройства и с их помощью найдены первичные основы волнового управления биосистемами на основе явления возврата Ферми-Паста-Улама и эпигено-лингвистических функций генетических молекул.
ЛИТЕРАТУРА
25. Гаряев П.П. Волновой геном. М., 1994. с.279.
26. Гаряев П.П. Кризис генетики и генетика кризиса. М., 1994. № 1- 6. с. 46- 49.
44. Гриневич Г.С. Праславянская письменность (результаты дешифровки). М., 1993. т1. с. 323.
45. Гурвич А.Г. Избранные труды. М., 1977.с. 351.
46. Инге-Вечтомов С.Г. Генетика с основами селекции. М., 1989. с. 404- 405.
47. Любищев А.А.О природе наследственных факторов. Пермь.,1925.с. 120.
48. Маковский М.М. Лингвистическая генетика. М., 1992.с. 190.
49. Налимов В.В. Спонтанность сознания. М.,1989. с.287.
51. Соломоник А. Семиотика и лингвистика. М., 1995.с. 346.
Научное издание
Петр Петрович Гаряев
Волновой генетический код
Редактор Н.Н.Кондракова
Художник В.Генерозова-Бугуева
Корректор Л.П.Горбачева
Подписано в печать 23.07.1997 г.
Формат 60? 84 1 /16 . Бумага мелованная. Печать офсетная
Уч.-изд. л. 7 .Усл. печ. л. 6,2. Усл. кр.-отт. 6.2
Тираж 2000 экз.
Отпечатано с готовых пленок
на полиграфпредприятии АО “Астра семь”
“ИЗДАТЦЕНТР” ЛР № 064326 от 27.11.95
129327, Москва, ул.Чичерина 2/9
Москва. Филипповский пер. 13.