Москва
Аннотация
Обсуждается проблема создания ДНК-волнового биокомпьютера, в котором будут использоваться ячейки памяти на генетических молекулах. В основу предлагаемой идеи положены экспериментальные и теоретические работы авторов, в которых a)доказана способность ДНК быть лазером, b)способность ДНК генерировать солитонные волны с памятью, c)обнаружено явление перехода локализованных фотонов в радиоволны, d)обнаружено явление спектрального запоминания ДНК локализованными фотонами, e)обнаружено явление переноса генетической информации в поляризационных модуляциях электромагнитного поля при переходе фотоны-радиоволны. На основе этих данных построена теория волновых генов, в которой предложено дуалистическое толкование генетической информации как единство вещественных и волновых кодирующих функций хромосом. Предложена гипотеза квантовой нелокальности генома высших организмов. Совокупность полученных результатов позволяет авторам предположить, что искусственный ДНК-компьютинг не может быть осуществлен в полной мере без учета перечисленных свойств генетического аппарата. Ключевой структурой ДНК-волнового компьютера будет являться феномен памяти ДНК на солитонах и на локализованных фотонах с элементами квантовой нелокальности такой памяти.
P.P. Gariaev (Ph.D., Dr.Sci.) Institute Control of Sciences Russian Academy of Sciences
G.G.Tertishniy (Ph.D.) Institute Control of Sciences Russian Academy of Sciences
E.A. Leonova Institute Control of Sciences Russian Academy of Sciences
A.V. Mologin Institute Control of Sciences Russian Academy of Sciences
Moscow
Summary
The problem of creation of the DNA-WAVE biocomputer is considered, in which one the storage locations on genetic molecules will be used. In the basis of tendered idea are trusted to experimental and theoretical activities of the writers, in which one a) to be demonstrated capacity of DNA to the laser, b) capacity of DNA to generate solitonic waves with memory, c) the phenomenon of transition of the localized photons in radio waves is revealed, d) the phenomenon of spectral storage of DNA by the localized photons is revealed, e) the transport phenomena of the genetic information in polarization modulations of an electromagnetic field is revealed at transition of a photon - radio wave. On the basis of these data the theory of wave genes is built, in which one the dualistic explanation of the genetic information as unity of material and wave encoding functions of the chromosomes is offered. The hypothesis quantum nonlocality of genome of higher organisms is offered. The set of the obtained outcomes allows the writers to suspect, that artificial DNA computing cannot be carried out to the full without the registration of listed properties of the genetic apparatus. Key pattern of the DNA-WAVE computer will be the phenomenon of DNA memory on solitons and on the localized photons with participation of quantum nonlocality of such memory.
В 1985г. одним из авторов были зафиксированы необычные колебательные режимы ДНК, рибосом и коллагена “in vitro” c использованием метода динамического лазерного светорассеяния. Недавно это было подтверждено нами и, в дополнение, обнаружен феномен трансформации лазерного света в радиоволны [27, 28]. Такая трансформация связана, вероятно, с эффектом квантовой нелокальности и регистрируется разработанным нами методом. Есть некоторые основания полагать, что генетический аппарат высших биосистем обладает способностью быть квантово нелокальным. Это дает возможность клеткам, тканям и организму находиться в суперкогерентном состоянии. Перечисленные результаты еще раз, но на более высоком уровне, подтверждают нашу теорию волновых генов [29, 30, 46]. Ключевым положением ее является то, что хромосомный аппарат биосистем функционирует одновременно как источник и приемник гено-знаковых лазерных, солитонных и голографических полей. Кроме того, хромосомный континуум многоклеточных организмов является неким подобием статико-динамичной мультиплексной пространственно-временной голографической решетки, в которой свернуто пространство-время организма. Но и этим не исчерпываются кодирующие возможности генетических структур. Последовательности нуклеотидов ДНК, образующие голографические и/или квази-голографические решетки, формируют еще и текстовые рече-подобные структуры, что существенно меняет наши представления о генетическом коде. Эволюция биосистем создала генетические "тексты" и геном-биокомпьютер как квази-разумный "субъект", на своем уровне "читающий и понимающий" эти тексты. Чрезвычайно важно для обоснования этой элементарной "разумности" генома, что естественные (не существенно на каком языке) человеческие тексты и генетические "тексты" имеют сходные математико-лингвистические и энтропийно-статистические характеристики. Это относится, в частности, к такому понятию как фрактальность распределения плотности частот встречаемости букв в естественных и генетических текстах (для генетических "текстов" буквы – это нуклеотиды) [1]. Ниже будут приведены полученные нами результаты о сходстве таких фракталов для генетических и естественных текстов.
Еще одно подтверждение лингвистической трактовки кодовых функций генома получено американскими исследователями [31]. Работая с “кодирующими” и “не кодирующими” последовательностями ДНК эукариот (в рамках старых представлений о генах), авторы пришли к выводу, противоречащему догме о том, что знаковые функции сосредоточены только в белок-шифрующих участках ДНК. Они применили метод статистического анализа естественных и музыкальных текстов, известный как закон Ципфа-Мандельброта, и принцип избыточности текстовой информации Шеннона, рассчитываемый как энтропия текстов. В результате оказалось, что не кодирующие районы ДНК более схожи с естественными языками, чем кодирующие, и что, возможно, не кодирующие последовательности генетических молекул являются основой для одного (или более) биологических языков. Авторы разработали также статистический алгоритм поиска кодирующих последовательностей ДНК, который выявил, что белок-кодирующие участки обладают существенно меньшими дальнодействующими корреляциями по сравнению с зонами, разделяющими эти участки. Распределение ДНК-последовательностей оказалось настолько сложным, что использованные методы переставали удовлетворительно работать уже на длинах, превышающих 103 пар оснований. Распределение Ципфа-Мандельброта для частот встречаемости “слов” с числом нуклеотидов от 3 до 8 показало большее соответствие естественному языку не кодирующих последовательностей по сравнению с кодирующими. Еще раз подчеркнем, что кодирование авторы понимают как запись информации об аминокислотной последовательности, и только. И в этом парадокс, заставивший их заявить, что не кодирующие регионы ДНК - это не просто “junk” (“мусор”), а структуры, предназначенные для каких-то целей с неясным пока назначением. Дальнодействующие корреляции в этих структурах авторам также непонятны, хотя и обнаружена нарастающая сложность не кодирующих последовательностей в эволюции биосистем. Эти данные полностью соответствует нашим идеям о том, что не кодирующие последовательности ДНК (а это 95%-99% генома) являются стратегическим информационным содержанием хромосом. Оно имеет, как нам представляется, материально-волновую природу, оно многомерно и выступает, в сущности, как ассоциативно-образная и лингвистико-волновая программа эмбриогенеза, смыслового продолжения и логического конца любой биосистемы. Интуитивно поняв это, авторы с ностальгической грустью прощаются со старой и хорошо послужившей моделью генетического кода, не предлагая, правда, ничего взамен.
Наши представления о генетическом коде должны существенно измениться, иначе мы никогда не сможем создать ДНК-компьютер. С этой целью мы постулировали механизм контекстно-волновых ориентаций рибосом для решения проблемы точного выбора аминокислоты [32]. Эта проблема возникла сразу при создании модели генетического кода как фактор неопределенности в выборе третьего нуклеотида в кодирующем аминокислоту триплете (wobble-гипотеза Ф.Крика). Для того чтобы понять, каким образом белок-синтезирующий аппарат клетки решает эту типично лингвистическую проблему снятия омонимической неопределенности необходимо ввести понятия фонового принципа, ассоциативно-голографической памяти генома и его квантовой нелокальности [28, 33, 38, 46]. Это означает, что геном может рассматриваться одновременно на уровне вещества и как идеальная (ментальная) структура, то есть как квантовый объект.
Универсальным информационным посредником в свертке-развертке знаковых регуляторных структур генома-биокомпьютера выступают эндогенные физические поля очень малой мощности. Эти поля продуцирует хромосомный аппарат, и они являются, быстрым волновым генетическим информационным каналом, соединяющим хромосомы отдельных клеток организма в целостный континуум, работающий как биокомпьютер. Кратко основные положения теории волновых генов (с учетом квантовой нелокальности генома) можно сформулировать следующим образом:
Эти положения необходимо учитывать в обсуждаемой здесь гипотетической модели биокомпьютера, работающего на генетических молекулах. Рассмотрим, как формируются “in vitro” конвертированные из фотонов радиоизлучения геноструктур (препараты жидких кристаллов ДНК). В наших экспериментах [28] мы получали так называемые локализованные или спутанные (entangle) когерентные фотоны с последующим пермиссивно-телепортационным превращением их в радиоволны. Этот процесс осуществлялся специально изготовленным одночастотным He-Ne лазером с мощностью излучения 2 мВт, длиной волны 632,8нм со стабильным резонатором, управляемым посредством электронного термостатирующего элемента [34]. При взаимодействии лазерного пучка с жидкими кристаллами ДНК (или с любыми другими объектами), лазер генерировал радиосигналы, различающиеся по характеру (спектру Фурье) в зависимости от типа исследуемых образцов и способов их приготовления. Одним из необходимых условий для генерации ДНК-знаковых биоактивных радиоволн является "трех зеркальная схема". В соответствии с ней зондируемый объект (ДНК) отражает лазерный пучок в обратно в резонатор лазера. Характерно, что специфические модуляции радиосигнала при этом полностью соответствуют изменению во времени двумерных спекл-картин рассеянного препаратами ДНК света.
В этих экспериментах мы получили первичную информацию о возможности долговременной записи биологически активной динамичной поляризационно-лазерно-радиоволновой генетической информации с препаратов ДНК на лазерных зеркалах, как самого лазера, так и на внешних лазерных зеркалах, не являющихся частями лазера (см. ниже). Мы предполагаем, что этот феномен связан с явлением локализации (сжатия) фотонных полей в системе коррелированных рассеивателей лазерных зеркал. В условиях плохого собственного поглощения излучения материалом таких рассеивателей внешнее световое поле способно удерживаться в системе в течение длительного времени без диссипации в другие формы энергии. Причина локализации связана с интерференцией многократно рассеянных волн. Внешний электромагнитный сигнал (в нашем случае это лазерный луч, промодулированный по поляризации препаратом ДНК, локализуется (“записывается”) в системе металл-содержащих неоднородностей лазерных зеркал. Этот сигнал в дальнейшем может быть “считан” без существенной потери информации уже в форме изоморфно (по отношению к фотонам) поляризованных радиоволн. В пользу этих соображений свидетельствуют теоретические исследования по сжатым состояниям локализованных фотонов [35-37].
Если такая "запись" на зеркалах
реальна, тогда содержащие атомы металлов
жидкокристаллические слои ДНК хромосомного
аппарата (аналоги зеркал) также можно
рассматривать как фрактальную среду накопления
локализованных фотонов, создающую когерентный
континуум с квантово-нелокально распределенной
поляризационно-радиоволновой гено-информацией.
Это в определенной мере соответствует ранее
высказанной нами идее квантовой нелокальности
генома, точнее одной из ее форм [32, 33, 38]. Возможно,
существует и другой механизм перехода квантов
света как солитонов в радиоволны. В работе
Тужински и соавторов [39] показана связь,
взаимодополняемость двух, казалось бы,
независимых теорий, в которых рассматриваются
две физические модели, объясняющие необычное
поведение биологических систем. Эти модели
предложены Гербертом Фрелихом и Александром
Давыдовым. Так называемые Давыдовские солитоны,
описывающие возбуждение, делокализацию и
движение электронов вдоль пептидных цепей
белковых молекул в форме уединенных волн
(солитонов) [40] дополняет известную модель
Фрелиха [41-44], развитую в нашей работе [45], о
возможности высоко поляризованного
(когерентного, лазероподобного) состояния
колеблющихся диполей информационных
биомакромолекул, диполей, возникающих при
Бозе-конденсации фононов электромагнитных волн
белков (![]() ), ДНК (
), ДНК (![]() ), мембран (
), мембран (![]() ). В работе Тужински и соавторов
Давыдовский гамильтониан трансформирован в
нормальные координаты, Фрелиховский
гамильтониан канонически трансформирован в
эквивалентную форму в рамках аппроксимации
Хартри-Фока. Авторы полагают, что модель
Гамильтониана способна связать обе теории,
которые математически эквивалентны. Кроме того,
обе модели дополняют друг друга физически.
Бозе-конденсация вибрационных мод биополимеров
соответствует распространению солитона волны
поляризации. И наоборот: солитонный транспорт
граничной энергии вдоль пептидной цепи
сопровождается Бозе-конденсацией решеточных
вибраций биоструктур. Отсюда следует, что
солитон порождает электромагнитное поле, а это,
возможно, один из механизмов эффекта, который мы
наблюдаем в экспериментах, когда осциллирующий
оптический солитон-бризер, отображающий
солитонные возбуждения ДНК, генерирует
оптико-резонансно усиленные радиоволны. Еще одна
мысль, привлекающая внимание: конверсия
эндогенных когерентных фотонов, генерируемых
хромосомами, в радиоволны в биосистеме может
происходить по "трехзеркальной" или
“многозеркальной” схеме на многочисленных
отражающих поверхностях мембран, аналогично
нашим модельным опытам. В этом случае клеточное
ядро (хромосомы) выступают как лазерный источник
света, а мембрана клеточного ядра и
цитоплазматические мембраны как полупрозрачные
зеркала. Доменные стенки жидкокристаллических
структур клетки также могут служить
“зеркалами” и одновременно являться при этом
зондируемыми объектами. В таком случае реальна
возможность “in vitro - in vivo” манипулировать
световыми лазерными потоками, которые
транспортируются сложнейшей сетью световодов
живой клетки и которые, вероятно, преобразуются
на клеточных структурах в радиоволны, несущие
информацию о структурно-метаболических
перестройках. Локализация и “запись” такого
рода фотонно-радиоволновой информации может
использоваться как основа для создания
искусственно биокомпьютерной памяти. В связи с
этим, в порядке научной полемики, можно
предложить создать ячейки памяти на жидких
кристаллах ДНК. Считывание информации с таких
ячеек осуществляется лазерным пучком в режимах,
разработанных нами. Как упоминалось выше,
первичные экспериментальные результаты в этом
направлении нами получены.
). В работе Тужински и соавторов
Давыдовский гамильтониан трансформирован в
нормальные координаты, Фрелиховский
гамильтониан канонически трансформирован в
эквивалентную форму в рамках аппроксимации
Хартри-Фока. Авторы полагают, что модель
Гамильтониана способна связать обе теории,
которые математически эквивалентны. Кроме того,
обе модели дополняют друг друга физически.
Бозе-конденсация вибрационных мод биополимеров
соответствует распространению солитона волны
поляризации. И наоборот: солитонный транспорт
граничной энергии вдоль пептидной цепи
сопровождается Бозе-конденсацией решеточных
вибраций биоструктур. Отсюда следует, что
солитон порождает электромагнитное поле, а это,
возможно, один из механизмов эффекта, который мы
наблюдаем в экспериментах, когда осциллирующий
оптический солитон-бризер, отображающий
солитонные возбуждения ДНК, генерирует
оптико-резонансно усиленные радиоволны. Еще одна
мысль, привлекающая внимание: конверсия
эндогенных когерентных фотонов, генерируемых
хромосомами, в радиоволны в биосистеме может
происходить по "трехзеркальной" или
“многозеркальной” схеме на многочисленных
отражающих поверхностях мембран, аналогично
нашим модельным опытам. В этом случае клеточное
ядро (хромосомы) выступают как лазерный источник
света, а мембрана клеточного ядра и
цитоплазматические мембраны как полупрозрачные
зеркала. Доменные стенки жидкокристаллических
структур клетки также могут служить
“зеркалами” и одновременно являться при этом
зондируемыми объектами. В таком случае реальна
возможность “in vitro - in vivo” манипулировать
световыми лазерными потоками, которые
транспортируются сложнейшей сетью световодов
живой клетки и которые, вероятно, преобразуются
на клеточных структурах в радиоволны, несущие
информацию о структурно-метаболических
перестройках. Локализация и “запись” такого
рода фотонно-радиоволновой информации может
использоваться как основа для создания
искусственно биокомпьютерной памяти. В связи с
этим, в порядке научной полемики, можно
предложить создать ячейки памяти на жидких
кристаллах ДНК. Считывание информации с таких
ячеек осуществляется лазерным пучком в режимах,
разработанных нами. Как упоминалось выше,
первичные экспериментальные результаты в этом
направлении нами получены.
Генетическая волновая информация с препаратов ДНК, записанная в поляризациях спутанных (entangled) фотонов, будучи квантово-нелокальной, переходит (разворачивается) в широкополосный радиоволновой спектр, изоморфный поляризациям фотонов. Именно модуляции поляризации фотонов-радиоволн оптически активными молекулами ДНК являются переносчиками квантово нелокальной морфогенетической и, шире, метаболической волновой информации. Поскольку фурье-образ радиоспектров существенно зависит от типа зондируемого вещества, мы предположили, что это явление может лечь в основу нового вида спектроскопии – поляризационно-лазерно-радиоволновой (ПЛР-спектроскопия) [28]. Фундаментальным фактом оказалось наблюдение, что фотонно-радиоволновые характеристики различных объектов (ПЛР-Фурье-спектры кристаллов, воды, металлов, ДНК и др.) запоминаются лазерными зеркалами и "живут" определенное время. Существенно, что эти "зеркальные спектры" (ПЛР-память) динамичны во времени, как и эквивалентные им спектры самого непосредственно зондируемого объекта. Эта весьма сложная и во многом непонятная нелинейная динамика c “памятью” имеет повторяющиеся во времени спектральные фигуры. Возможно, и здесь реализуется явление возврата Ферми-Паста-Улама солитонного типа, которое мы уже наблюдали в случае нелинейной динамики ДНК при светорассеянии на ее гелях [29]. Характерные повторы спектральных образов индуцированных радиоволновых излучений препарата ДНК приведены на Рис.4. Это первый пример того, что относительно статичная, многослойная среда записи (лазерные зеркала) хранит в себе динамичную спектральную информацию об объектах записи. Обнаруженные феномены могут дать реальные основания для разработки принципиально нового типа видеозаписи, а также создания нового кинематографа.
При дальнейших исследованиях обнаружилась высокая биологическая (генетическая) активность радиоволн, генерируемых в таких условиях препаратами ДНК. С помощью таких ДНК-излучений нам удалось вызвать сверхбыстрое развитие клубней картофеля вне почвы (удлинение ростков до 1 см в сутки) и резкие изменения его морфогенеза с образованием небольших клубней не на корневищах, а на стеблях. Эти же излучения оказались способными вызвать статистически достоверное "оживление" старых и мертвых семян Arabidopsis thaliana, взятых из зоны Чернобыля в 1987 году. Контрольные облучения поляризованными радиоволнами, не содержащими информации о ДНК биологически не активны [33]. В этой серии экспериментов мы получили еще одно доказательство возможности существования генетической информации в форме поляризационно-лазерно-радиоволнового физического поля. О такой возможности вот уже около 70 лет говорят и спорят биологи.
Мы предполагаем, что главный
информационный канал в этих экспериментах с ДНК
– биознаковые связанные (“спутанные”)
модуляции поляризаций фотонов и радиоволн при
переходах "фотоны ![]() радиоволны" с сохранением информационной
связи между ними в рамках одного из вариантов
квантовой нелокальности (см. ниже). По этой
причине совершенно в ином ракурсе видится хорошо
известный факт, что информационные
биомакромолекулы – ДНК, РНК и Белки – обладают
ярко выраженной способностью к дисперсии
оптического вращения и круговому дихроизму. Это
обнаруживается в характеристическом (зависимом
от длины волны и от свойств образца) вращении
электромагнитных векторов фотонов и разном
поглощении фотонов образцами в зависимости от
типа образца и от того, вправо или влево
закручиваются электрический и магнитный вектора
поля фотонов. Низкомолекулярные компоненты
биосистем, такие как сахара, нуклеотиды,
аминокислоты, порфирины и другие вещества
обладают той же способностью. До сих пор в этом не
видели биологического смысла. Теперь же феномен
оптической активности может быть понят как база
для получения организмом неисчерпаемой
информации о собственном метаболизме.
Информация считывается эндогенными лазерными
излучениями хромосом, переходящими в
регуляторное ("смысловое") радиоизлучение
генома-биокомпьютера. Снимается и противоречие
между длинами радиоволн таких превращенных
излучений и размерами организмов, клеток и
субклеточных структур. В этой ситуации смысловые
резонансы в пространстве-времени биосистем
происходят не на уровне длин волн, а на уровне
частот и углов поворотов поляризационных мод.
Вместе с тем, это и база для искусственного
лазерно-радиоволнового "in vitro - in vivo"
сканирования организма и его компонентов, как
нового вида спектроскопии [28].
радиоволны" с сохранением информационной
связи между ними в рамках одного из вариантов
квантовой нелокальности (см. ниже). По этой
причине совершенно в ином ракурсе видится хорошо
известный факт, что информационные
биомакромолекулы – ДНК, РНК и Белки – обладают
ярко выраженной способностью к дисперсии
оптического вращения и круговому дихроизму. Это
обнаруживается в характеристическом (зависимом
от длины волны и от свойств образца) вращении
электромагнитных векторов фотонов и разном
поглощении фотонов образцами в зависимости от
типа образца и от того, вправо или влево
закручиваются электрический и магнитный вектора
поля фотонов. Низкомолекулярные компоненты
биосистем, такие как сахара, нуклеотиды,
аминокислоты, порфирины и другие вещества
обладают той же способностью. До сих пор в этом не
видели биологического смысла. Теперь же феномен
оптической активности может быть понят как база
для получения организмом неисчерпаемой
информации о собственном метаболизме.
Информация считывается эндогенными лазерными
излучениями хромосом, переходящими в
регуляторное ("смысловое") радиоизлучение
генома-биокомпьютера. Снимается и противоречие
между длинами радиоволн таких превращенных
излучений и размерами организмов, клеток и
субклеточных структур. В этой ситуации смысловые
резонансы в пространстве-времени биосистем
происходят не на уровне длин волн, а на уровне
частот и углов поворотов поляризационных мод.
Вместе с тем, это и база для искусственного
лазерно-радиоволнового "in vitro - in vivo"
сканирования организма и его компонентов, как
нового вида спектроскопии [28].
Представляется, что обсуждаемый случай квантовой нелокальности хромосом, как проявление нелокальности генетической информации, является частным случаем. Нелокальность генетической информации в высшей степени характерна для многоклеточных организмов и распределена по уровням.
1-й уровень – организменный. Нелокальность здесь выражается в способности к регенерации, например у червей планарий. После разрезания таких червей любая часть их тела дает при регенерации целый организм. Иными словами, в этом случае отсутствует привязка общего пула генетической информации к какой-то части биосистемы. То же относится к вегетативному размножению растений.
2-й уровень – клеточный. Из каждой клетки, а не только из зиготы, можно вырастить целый организм. Для животных биосистем это затруднено, но возможно. Каждая клетка – потенциальный континуум организма.
3-й уровень – клеточно-ядерный. Энуклеация ядер из соматических и половых клеток с последующим введением в них других ядер не препятствует развитию нормального организма. Клонирование такого рода уже осуществляют на высших биосистемах, например, на овцах. Каждое клеточное ядро – также потенциальный континуум биосистемы. Локализации генетических потенций на каких-то отдельных клетках нет.
4-й уровень – молекулярный: рибосома "читает" информационную РНК не только по отдельным кодонам, но и всю ее целиком с учетом контекста, то есть нелокально, континуально.
5-й уровень – хромосомно-голографический. Геном обладает голографической памятью [23], а это типично распределенная (нелокальная) ассоциативная память. На этом и последующих уровнях нелокальность приобретает новое качество, дуалистический вещественно-волновой характер, поскольку голограммы как вещество "прочитываются" электромагнитными и/или акустическими полями, выносящими гено-волновую информацию за пределы вещества хромосом. На сцену выходит физическое поле или поля, как калибровочные, размечающие будущее пространство-время организма. Сюда же относится, видимо, голографическая память коры головного мозга, задающая ментальные, смысловые и образные пространства, калибрующие (векторизующие) потенциальные действия высших биосистем. А это уже высший, социогенетический ареал работы генома.
6-й уровень – квантовая нелокальность хромосомного континуума. До 6-го уровня нелокальность генетической информации реализуется в пространстве и времени организма. При этом время и пространство константны и не имеют градиентов (искривлений). 6-й уровень имеет особый характер и новое качество. Оно проявляется в рамках одной из форм квантовой нелокальности, а именно пермиссивной, постулируемой в нашей работе [28]. В этом случае нелокальность реализуется как по пространству биосистемы, так и по ее собственным, “сжимаемым” до нуля, пространстве-времени. Мгновенно распространяемые такими способами гено-волновые программы, изоморфные вещественным, работают в организме "здесь и там одновременно", поэтому утрачивает смысл семантическая конструкция "сначала и потом". И это стратегический фактор, необычайно важное для многоклеточных биосистем эволюционное достижение. Миллиарды клеток организма должны "знать" друг о друге если не все, то очень многое (стратегическое), причем мгновенно. Без явления “волновой информационной мгновенности” гигантский многоклеточный континуум высших биосистем не способен целостно координировать метаболизм, свои физиологические и другие функции. Межклеточная диффузия сигнальных веществ и нервные процессы слишком инертны для этого. Даже если допустить, что в межклеточной передаче участвуют знаковые электромагнитные поля со световыми скоростями, что достаточно вероятно, то и этого недостаточно. Необходим механизм именно квантово-радиоволновой нелокальности, и он применим к генетическому аппарату, который может выступать как мгновенно распределенный развернутый квантовый (волновой) объект, изоморфный свернутой вещественной информации хромосомного континуума. Используя такую нелокальность, генетический аппарат высших биосистем создает удивительное явление, когда в определенных знаковых ситуациях в “схлопнутом” пространстве-времени биосистемы "здесь и там", "сначала и потом" работают как неразрывность, обеспечивающая организмам суперкогерентность, информационную сверхизбыточность, сверхинформированность, связность и, как итог, целостность (выживаемость). Проявлением этого, например, служит способность к регенерации органов и тканей у низших организмов (гидры, черви, амфибии, ящерицы, ракообразные), способность, которая в значительной степени утрачена человеком. Но ее можно активировать, учитывая развиваемые нами принципы волновой самоорганизации биосистем. Иллюстрацией этого служит первое в мире успешное приживление имплантированных слепому человеку донорских тканей с восстановлением зрения, осуществленное Э.Р.Мулдашевым [47]. В основу идеологии такой хирургической операции и регенеративных процессов были положены наши исследования [29, 30, 46], в том числе совместные с Э.Р.Мулдашевым [28, 48]. Вместе с тем, теоретико-экспериментальные исследования в этой области знания все еще носят начальный характер и нуждаются в дальнейшем физико-математическом осмыслении и развитии.
В применении к биокомпьютерам аналоги таких нелокальных процессов и ПЛР-память, возможно, станут основой в развитии вычислительной техники вообще. Это будет полная смена элементной базы и, в некотором смысле, повторение пройденного на пути развития вычислительной техники на совершенно ином, качественно более высоком, уровне в ряду: аналоговый>цифровой>“образный”. Последний и будет являться смысловым нелокальным волновым компьютером на ДНК.
Однако логика развития исследований в этой области поначалу идет в ином направлении. Молекулы ДНК стали использовать как чисто вещественные “параллельно вычисляющие” структуры. Это началось в 1994 году, когда Леонард Адлеман, профессор вычислительных наук из Университета Южной Калифорнии, предложил алгоритм использования ДНК для решения одной из версий “задачи коммивояжера” [49]. Эта задача является одним из выражений так называемой проблемы Гамильтониановского Пути в тяжелых математических задачах (Hamiltonian Path Problem или HPP), и она связана с перебором огромного числа вариантов возможных решений для получения оптимального. Адлеман с помощью “ДНК-компьютинга” решил задачу для 7 городов и 13 дорог между ними, когда необходимо проложить кратчайший маршрут однократного посещения каждого этих городов. Потребовалась всего неделя для получения ответа, в то время как традиционным компьютерам понадобилось бы несколько лет. При этом было использовано фундаментальное явление, свойственное молекулам ДНК – способность ее одиночных цепей к комплементарным взаимоузнаваниям. Это явление заключается в том, что любые фрагменты каждой из двух цепочек ДНК находят в растворе (или в составе хромосом живой клетки) только собственные, в некотором смысле зеркальные, половинки и образуют нормальную двойную спираль. Этот феномен является одним из проявлений общего свойства высокоорганизованных биоструктур и полимерных молекулярно-надмолекулярных образований к самосборке. Так in vitro - in vivo самособираются рибосомы, мембраны, хромосомы, вирусы и фаги. В том числе и однонитевые ДНК. Успешность и быстрота спонтанных поисков половинками ДНК друг друга, как акта самоорганизации (самосборки) и обеспечили высокую скорость перебора вариантов в пределах “задачи коммивояжера”. Причины быстрых и точных взаимоузнаваний половинок ДНК до недавнего времени были неизвестны. А это необычайно важно для реального создания ДНК-компьютера, и об этом речь пойдет ниже. Несколько подробнее о модели Адлемана, поскольку его и наша логики принципиально различаются. Как мы (и не только) полагаем, путь, который выбрал Адлеман и его многочисленные последователи, используя ДНК как “вычислительную” структуру, неправильно ими оценивается как некий ДНК-компьютинг. Дэвид Гиффорд, один из крупных авторитетов в компьютинге, первым поддержавший Адлемана, сказал, что “это не молекулярный компьютер”, и что эта техника “..может только решать некоторые виды комбинаторных проблем, это не универсальный или программируемый компьютер типа IBM PC” [50]. Чтобы понять, почему правы мы и Гиффорд, коротко рассмотрим метод Адлемана. Он обозначил каждый город как отрезок однотяжной ДНК длиной в 20 оснований (баз) со случайными последовательностями. Дороги между каждыми двумя городами были представлены как отрезки комплементарных однотяжных ДНК в 20 баз, которые перекрывают половины путей между городами. При этом соблюдается каноническое правило спаривания оснований в двутяжных ДНК: Аденин-Тимин, Гуанин-Цитозин. Путь между 7 городами начинается с фрагмента двутяжной ДНК, которая соединяет какие-либо два города. Важно, что фрагментов ДНК, обозначающих какой-то один город, может быть больше чем один. Затем более 100 миллиардов радиоактивно меченых “ДНК-городов” и “ДНК-путей” были перемешаны в пробирке и размножены ферментативной ДНК-амплификацией. На этом, как считает Адлеман, “ДНК-компьютинг” заканчивается. Далее, чтобы получить ответ – оптимальный путь (определенные фракции ДНК), реакционную смесь с “ответом” электрофоретически разделяли с тем, чтобы получить все пути, идущие от “старта” до “конца”. Затем выделяли те пути, которые только раз проходили через 7 городов; выделяли пути между 7 разными городами. И если обнаруживали фракции “ДНК-путей” после этого этапа, то они считались наиболее оптимальными (“победителями”). В этом и было “решение” задачи коммивояжера. В процессе нахождения такого “решения” были задействованы миллиарды параллельных быстро происходящих комплементарных спонтанных (не программируемых человеком) актов “узнаваний” однотяжных ДНК и миллиарды спонтанных энзиматических репликаций этих молекул. При этом с малой затратой времени и энергии образуется нечто вроде “генетического супа”. Такая скорость и точность молекулярных процессов немыслима для эквивалентных операций в цифровых электронных компьютерах, использующих детерминистические вектора обработки информации. В случае “ДНК-компьютинга”, как считают, используются не детерминистические акты обработки больших параллельных массивов цифр-букв (4-х нуклеотидов ДНК). Итак, Алгоритм решения Гамильтониановского пути по Адлеману таков:
Молекулярно-биологические этапы получения решения сводятся к следующим операциям:
а) синтез однотяжных ДНК,
б) разделение их по длине с вычленением 20-базовых ДНК,
в) смешивание их в пробирках,
г) выделение нитей ДНК с известными последовательностями,
д) выделение реассоциацией комплементарных двутяжных ДНК,
е) PCR-амплификация (размножение) ДНК,
ж) разрезание ДНК рестриктазами,
з) лигирование ДНК, комплементарных по “липким” концам,
и) определение присутствия или отсутствия меченых ДНК в тестовых пробирках.
Какова эффективность работы такой системы “вычислений? В то время как существующие цифровые компьютеры производят 109 операций на Джоуль, “ДНК-компьютер” может делать 2·1019 операций на Джоуль, то есть в 1010 более эффективно. Плотность информации в ДНК - 1 бит/nm3, а в существующих компьютерах 1012 nm3 содержат 1 бит”[50].
И все-таки, является ли такая работа ДНК в рамках такой методологии работой компъютера? Нет. В этом варианте, в контролируемых условиях, спонтанно, в параллельных режимах нарабатывается огромное количество “ДНК-путей”. В том числе и правильных (оптимальных). Далее начинается собственно компьютинг, но он осуществляется людьми. Осмысленное выделение фракций ДНК – это и есть процесс получения решения задачи коммивояжера. В роли компьютера здесь выступает человек, его ментальное участие есть условие получение ответа. Но это не участие в программировании ДНК, что сближало бы такую работу с известным цифровым компьютингом. ДНК уже сама по себе “запрограммирована” на комплементарность в ходе эволюции живых систем. Однонитевые ДНК изначально способны к взаимоузнаваниям. Принципиально, что комплементарность Аденин-Тимин, Гуанин-Цитозин только на последних этапах обеспечивается близко действующими водородными связями между азотистыми основаниями. Предварительные прицельные “наводки” как между однонитевыми ДНК, так и между тРНК-мРНК антигеном-антителом и т.д. осуществляются на уровне дальних волновых взаимодействий (“узнаваний”). Эту способность ДНК можно назвать элементарной потенцией к распознанию образов, а следовательно, компьютингу. Но это явление другого рода, а именно, ДНК-волновой компьютинг. Принципиальное отличие ДНК-волнового компьютинга от электронно-цифрового в том, что он оперирует образами и квази-речевыми построениями [1, 30]. Такой биокомпьютер работает не с цифрами, как эквивалентами богатства (такого, например, как валюта), а с самим богатством. Как упоминалось, “задачи коммивояжера” успешно и спонтанно, без ментального участия человека, решаются в таких актах самосборки in vitro - in vivo как биогенез рибосом, вирусов, мембран, полисубъединичных белков, а также в процессах самоорганизации хромосомного аппарата после митоза и мейоза. Кроме того, эти механизмы используются живой клеткой при нахождении путей сближения в реакциях антиген-антитело, тРНК-мРНК, белок-рецептор и т.д. В этих актах достигается быстрый перебор и нахождение оптимальных волновых векторов самоорганизации биосистем, высшим проявлением которой служит биоморфогенез.
Параллелизм и амплификация рестриктных фрагментов ДНК с размножением множества “решений” в модели “компьютинга” Адлемана можно также рассматривать как образец искусственной нелокальности создаваемых в пространстве реакционных пробирок “ДНК-смысловых” ареалов, поскольку отсутствует пространственная и временная привязка точного “решения” задачи коммивояжера. Локальность здесь возникает только после принятия истинного решения в локальной голове человека после отбора им определенных фракций “ДНК-победителей”.
Правильное и эффективное использование ДНК, как основного информационного элемента потенциального биокомпьютера, немыслимо без понимания новых функций генетических молекул в биосистемах. Казалось бы, относительно роли ДНК все ясно – давно открыт генетический код, имеется с полдюжины нобелевских лауреатов. Вроде бы, налицо и успехи в генной инженерии. Однако в последние годы выяснилось, что все далеко не так безоблачно. Фактически сейчас генетика и эмбриология вышли на новый уровень, когда полученных знаний о ДНК, как носителя известного триплетного кода белков, оказывается недостаточно. Как и десятилетия назад мы не знаем главного - каким образом записана информация о строении нашего тела в хромосомах, как она считывается. Общепринятая модель генетического кода - лишь слабое приближение к пониманию программ создания организма. Уже то, что такая модель предполагает большую часть ДНК в хромосомах “мусорной”, не выполняющими никакой роли, ставит под сомнение ее правильность. Именно эта "не кодирующая" часть хромосомного материала требует иного мышления, особенно в попытках создать ДНК-компьютер, не говоря уже о нашем желании понять феномен возникновения Жизни.
Напомним, что хромосомный аппарат, как система записывающая, сохраняющая, изменяющая и транслирующая генетическую информацию, может рассматриваться одновременно на уровне Вещества и на уровне достаточно хорошо изученных Физических Полей, которыми, как носителями генетической и общерегуляторной информации, оперирует континуум молекул ДНК, РНК и белков. Здесь реализуются, как показали наши исследования, неизвестные ранее виды памяти (солитонная, голографическая, поляризационная), и при этом молекулы ДНК могут работать как биолазеры и как среда записи лазерного сигнала [21, 29]. Кроме того, мы обнаружили, что ДНК способна излучать индуцированное лазером широкополосное электромагнитное радиоволновое поле (см. выше). Рассматриваемый с таких позиций генетический код будет существенно иным по сравнению с “канонической”, но неточной моделью. Прежняя модель генетического кода может объяснить только механизмы биосинтеза белков живых организмов. Поэтому она является системой трактовок лишь начальных звеньев в сложной иерархической цепочке вещественных и волновых голографических, семиотико-семантических, в общем случае образных, кодирующих и декодирующих функций хромосом. Молекулы ДНК, как гено-знаковый континуум любой биосистемы, способен к формированию прообразов биоструктур и организма в целом как реестр динамичных, сменяющих друг друга “волновых копий” или “матриц”, изоморфных архитектонике организмов. Этот континуум является разметочным, калибровочным полем для построения биосистемы. В этом плане механизм быстрого и точного взаимоузнавания однотяжных ДНК, тот механизм, которым воспользовался Адлеман для решения “задачи коммивояжера” - лишь один из способов самоорганизации биосистем. Взаимоузнавание, в частности, происходит потому, что в молекулах ДНК зарождаются особые сверхустойчивые акустико-электромагнитные волны, солитоны, некоторые разновидности которых можно трактовать в рамках открытого в 1949г. явления возврата Ферми-Паста-Улама (ФПУ). Такие солитоны ДНК обладают памятью, свойственной явлению ФПУ-возврата. Она выражается в том, что нелинейные системы способны помнить начальные моды возбуждений и периодически к ним “возвращаться”. Напомним, что жидкие кристаллы ДНК в составе хромосом – это типичная нелинейная система. Другой тип памяти ДНК-континуума в организме – квазиголографическая, она же и фрактальная, поскольку любая голограмма есть фрактал. Такая память – одно из проявлений нелокальности генома (см. выше), и она связана с фундаментальным свойством биосистем - восстанавливать целое из своей части. Это свойство хорошо известно (черенкование растений, регенерация хвоста у ящериц, регенерация целого организма из яйцеклетки). Наиболее развитая форма такой памяти – голографическая (ассоциативная) память коры головного мозга, то есть нейронов. Все эти результаты приводятся здесь только потому, что бесперспективно рассуждать о ДНК-компьютере, даже решив с помощью молекул ДНК “задачу коммивояжера”, если не учитывать новую логику в понимании волновых знаковых, кодирующих биофункций ДНК.
Уединенные волны (солитоны) ДНК, пробегающие по ее длине, могут выступать в качестве "субъектов чтения" знаковых структур генома. Такую роль выполняют волны крутильных колебаний нуклеотидов в однотяжных участках ДНК, а также в РНК [11]. Знаковая колебательная динамика таких кручений нуклеотидов является, вероятно, одной из многих нелинейно-динамических семиотических образований генома. Что касается термина “тексты ДНК”, который был взят взаймы у лингвистов для метафорического употребления, то оказывается эта текстовая структура ДНК действительно сродни человеческой речи. Наши математико-лингвистические исследования [1, 5, 7] показали, что такой ключевой параметр, как фрактальность, един для ДНК и человеческой речи. Это видно при сравнении Рис. 1а, на котором дана матрица плотности хаотически игрового представления некоторой проекции текста на английском языке, и Рис. 1б, на котором приведена аналогичная матрица нуклеотидной последовательности, кодирующей первичную структуру белка казеина. Такие наблюдения коррелируют с ранними работами в этой области (см., напр., работы Н.Хомского по универсальным грамматикам или монографию. М.М.Маковского “Лингвистическая генетика” (1992г.)). Используя эти теоретические разработки и собственные данные по физикохимии ДНК, нам удалось экспериментально доказать возможность свертки генетической информации в форме солитонных волновых пакетов, описываемых физико-математическим формализмом явления возврата Ферми-Паста-Улама (ФПУ). Такие волновые пакеты с искусственно введенной в них биоинформацией, генерируемые разработанными нами ФПУ-радиоэлектронными устройствами, способны входить в резонансный информационный контакт с генетическим аппаратом животных, растений и, вероятно, человека с последующим резким и направляемым изменением их обмена веществ. Оказалось, что и само вещество наследственности - ДНК - является генератором ФПУ-солитонных акустико-электромагнитных полей. Именно поэтому ФПУ-генераторы способны вводить волновую информацию в хромосомы по электромагнитным резонансным механизмам. Эффективность ФПУ-генераторов на порядки возрастает, если на практике использовать феномен математической общности фрактальной структуры ДНК-“текстов” и человеческой речи [1, 30]. Грамматика генетических текстов является, вероятно, частным случаем универсальных грамматик всех языков людей. Поэтому и реализуются физико-смысловые резонансы солитонных структур ДНК и искусственных знаковых ФПУ-солитонных полей, как аналогов естественных ФПУ-хромосомных полей. Вводя определенные кодовые вербальные команды через генератор ФПУ в генетический аппарат радиационно поврежденных семян пшеницы и ячменя, нам удалось достоверно уменьшить число хромосомных аберраций, то есть фактически блокировать поражающее действие рентгеновского облучения. Более того, выяснилось, что возможна превентивная защита генома растений от жесткого рентгеновского излучения с помощью адекватных волновых команд. Контрольные эксперименты, с хаотическими вербальными построениями (командами), введенными через ФПУ-устройства в геном биосистем, показали, что такие команды никак не влияют на хромосомы. Эти эффекты предсказаны и проверены на основании теории волновых генов и с использованием математических компьютерных моделей, имитирующих “чтение” солитонами на ДНК генотекстов и ретрансляцию этих текстов в другие клетки и ткани [1-26]. На Рис. 2 и 3 представлены результаты численного моделирования динамики конформационных возмущений ДНК [11], показывающие зависимость поведения уединённой (солитоноподобной) волны от последовательности нуклеотидов ДНК, на которой эта волна была запущена. Другие наши физико-математические модели и эксперименты обосновывают т.н. “антенный эффект” при возбуждении электромагнитными полями выделенных коллективных мод макромолекул ДНК. Это прямо связано с теорией волновых генов, экспериментами по двухфотонной накачке геноструктур in vitro с последующим лазерным излучением ДНК, а также согласуется с результатами по запоминанию жидкими кристаллами ДНК инфракрасного импульсного лазерного сигнала [29].
Вернемся к гипотетическому биокомпьютеру, использующему вещественно-волновые знаковые функции ДНК. Ясно, что при его разработке необходимо использовать не только и не столько результаты эксперимента Адлемана и его последователей. Чтобы реализовать свои возможности in vitro, ДНК и/или хромомомы должны находиться в естественной для них среде - в водном растворе, имитирующем кариоплазму, и в жидкокристаллическом состоянии. Истинные волновые управляющие, в том числе и компьютерные, возможности геноструктур могут быть выявлены в условиях, максимально приближенных к тем, которые имеются в живой клетке. В пределе компьютер на ДНК - это и есть живая клетка. Искусственный аналог клетки пока невозможен. Сейчас мы можем делать только какие-то модели приближения к волновым знаковым состояниям ДНК в клетке, как это было сделано нами в отношении записи ДНК-волновой информации на лазерных зеркалах и регенерации радиационно поврежденных семян ДНК-радиоволнами (см. выше). Далее необходимо начать практическое использование волновых типов памяти геноструктур и для этого пытаться конструировать ячейки памяти, работающие на явлении ФПУ-резонансов и/или на способности записывать голограммы, а также на явлении записи поляризационно-лазерно-радиоволновой ДНК-информации на локализованных (сжатых, спутанных) фотонах. Такая память будет на многие порядки по объему, быстродействию, “интеллектуальности” превосходить память существующих магнитных, оптических дисков и голографических запоминающих установок. Вторая принципиальная возможность связана с перечисленными типами памяти, но многократно усиливается способностью хромосом быть лазероактивной средой. Препараты хромосом выступают в таком варианте одновременно и как ячейки памяти, и как лазеры, считывающие собственную (а также наведенную) голографическую, ФПУ-память и память на локализованных фотонах. И наконец, последняя из достижимых в настоящее время целей - использование квази-речевых характеристик ДНК. Можно создавать такие ДНК-лазеры, которые будут высвечивать и “озвучивать” как естественные генотексты, так и искусственные (синтезированные человеком) знаковые последовательности полинуклеотидов, имитирующие естественные квази-речевые генопрограммы. Однако это весьма опасный путь и необходима система запретов на искусственные волновые гены. Такой способ работы с потенциальными ДНК-компьютерами означает вхождение в новые семиотические ареалы генома человека, вообще всей биосферы, ареалы, которые Природа использовала для создания человека. Эта мысль вполне обоснована, если учесть теоретические работы по коллективной симметрии генетического кода, проводимые школой Эйгена в Институте Макса Планка в Германии. Исследования школы Эйгена показывают, что ключевая часть информации, записанная и записываемая как квази-речь в хромосомах всех организмов нашей планеты, носит искусственный характер. Наши данные о том, что хромосомный континуум и ДНК любой биосистемы является неким подобием антенны, открытой во вне для приема дополнительной (возможно, экзобиологической) Информации, подтверждают сказанное [52]. Можно думать, что геном организмов Земли, по крайней мере частично, является полигоном для смысловых Экзобиологических влияний, и в этом плане существенно, что мы нашли первичные подходы к вхождению в этот семиотико-семантический ареал. Основываясь на сказанном, можно предсказать, что открываются следующие перспективы знаковых манипуляций с геноструктурами, как основным субстратом биокомпьютеров: а) создание искусственной памяти на генетических молекулах, обладающей поистине фантастическим объемом и быстродействием, б) Создание биокомпьютера на ДНК, основанного на волновых принципах и сравнимого по способам обработки информации и функциональным возможностям с человеческим мозгом, в) осуществление дистантного управления ключевыми информационными процессами в биосистемах через искусственные биокомпьютеры (лечение рака, СПИДа, генетических уродств, управление социогенетическими процессами и, в конечном итоге, увеличение времени жизни человека), г) активная защита от деструктивных волновых влияний через обнаруженный информационно-волновой канал, д) устанавление экзобиологических контактов.
Подводя итог, спросим - что остается от логики постановочных экспериментов с ДНК, которую предлагают Адлеман и другие исследователи в области ДНК-компьютинга? Эта логика уязвима, поскольку основана на упрощенных представлениях о знаковой работе хромосом, как только вещественного субстрата. Волновые знаковые функции геноструктур не берутся в расчет. Это неизбежно приводит в тупик в попытках использовать одномерное мышление относительно ДНК при создании биокомпьютера. В действительности такой компьютер должен имитировать функции генома в оперировании волновой информацией - создавать образы, в том числе и квази-речевые, распознавать их, манипулировать ими как командными. ДНК-компьютерные волновые знаковые структуры будут обладать огромной биологической, а может быть, и ментальной активностью. Если принять эти идеи, то необходимо иное стратегическое распределение финансирования в генетике, эмбриологии и генной инженерии, а также в ДНК-компьютинге. ДНК-волновые компьютеры будут способны управлять суперсложными процессами, реально сравнимыми с метаболизмом и мышлением. Это тем более вероятно, поскольку геном, как нам представляется, использует эффекты квантовой нелокальности.
В 1935г. А.Эйнштейн, Б.Подольский и Н.Розен [53] высказали идею, суть которой сводится к следующему. Квантовый объект, например два связанных фотона, в процессе разделения и разлета сохраняют некое подобие информационной связи (эффект "спутывания", "связывания" (entangle)). При этом квантовое состояние одного, например поляризация или спин, может мгновенно (за нулевое время) передаваться на другой фотон, который при этом становится аналогом первого, а первый коллапсирует, исчезает. И наоборот. Расстояние между фотонами может быть любым. Эта чисто теоретическая идея была названа эффектом, парадоксом или каналом Эйнштейна-Подольского-Розена (ЭПР). Как синоним этого феномена принят также термин "Квантовая Нелокальность" (Quantum NonLocality). Он подчеркивает мгновенную распределенность, нелокальность в пространстве состояний связанных по квантовым состояниям элементарных частиц. Казалось бы, нарушается принцип причинности - следствие и причина не разделены временем, если понимать время как способ организации последовательности событий. Поэтому Эйнштейн и соавторы, не располагая тогда знаниями о сложной структуре времени (к примеру, о его фрактальности), оценивали свою чисто теоретическую, но, тем не менее, жестко формализованную, модель как неприложимую к практике, эксперименту. Это состояние противоречия теории и видимой физической реальности длилось около 30 лет. Затем Д.Белл [54] развил идею ЭПР на современном уровне. Активное участие в этом приняли также Ч.Беннет и соавторы [55]. Главная трудность состояла в том, что необходимо было в теоретических построениях не нарушать фундаментальный принцип квантовой механики, выведенный Гейзенбергом для дуалистичного вещественно-волнового понимания квантовых объектов. Это принцип неопределенности о невозможности правильного одновременного измерения свойств, например фотона, как волны и как элементарной частицы. Эта проблема была снята после экспериментального доказательства возможности существования "спутанного" (entangle) состояния элементарных частиц. Возможно, такая "спутанность" - это элементарное основание передачи волновой генетической информации между клетками организма. Не существует запрета рассматривать живые клетки и ДНК как континуум элементарных частиц. В спутанном состоянии, обе рассматриваемые элементарные частицы остаются частью одной и той же квантовой системы так, что все, что Вы делаете на одной из них, предсказуемо влияет на другую. Беннет и его коллеги считают, что спутанные частицы при их разделении в пространстве могут служить в качестве взаимных "переносчиков" своих состояний и, следовательно, взаимной информации, поскольку любое измененное состояние частицы - это уже информация. Для экспериментальной реализации ЭПР-канала, необходимо было сосуществование трех фотонов - спутанного и двух разлетающихся, что и было осуществлено работами двух групп исследователей - венской, возглавляемой Антоном Цойлингером, и римской, под руководством Франческо Де Мартини. Опыты группы Цойлингера [56] и Де Мартини доказали выполнимость принципов ЭПР на практике для передачи через световоды состояний поляризации между двумя фотонами посредством третьего на расстояниях до 10 километров. После этого открытия в ведущих странах осуществляются мощные программы по применению этого эффекта для создания квантовых оптических компьютеров и разработки квантовой криптографии, где носителями информации и системой ее защиты будут фотоны. Скорость их работы и объемы информации будут на десятки порядков превосходить таковые у существующих компьютеров (особенно, если это лазерные фотоны жидких кристаллов хромосом). Идея использования явления Квантовой Нелокальности биосистемами весьма привлекательна и в мировоззренческом, и в практическом планах, в том числе и для создания ДНК-компьютеров. Она хорошо соответствует нашим данным о волновом знаковом назначении гено-информационно-метаболических и ментальных ареалов биосистем. В этом смысле первую, но довольно слабую, попытку разобраться в приложимости концепции ЭПР к биосистемам сделали несколько ранее [57]. Указанная работа констатирует, что восприятие действительности организмами базируется на ином и, в определенном смысле, более эффективном принципе, чем тот, что используется более формальными процедурами в науке. Этот принцип, по мысли авторов, в некоторых условиях реализуется в "не физичных" интеркоммуникационных знаковых взаимодействиях не статистического характера между пространственно разделенными биосистемами, то есть в телепатии. Почему "не физичных" и какое к этому имеет ЭПР, остается неясным.
Еще раз поставим вопрос, но более узко, и не затрагивая преждевременно проблему телепатии, - участвует ли явление квантовой нелокальности в работе генетического аппарата высших биосистем? Если да, то каким образом? Ясно, что даже предположения здесь будут носить сугубо предварительный характер. Однако, необходимость в рабочих гипотезах здесь уже назрела. В волновых версиях работы генома [29, 30] ЭПР-эффект является желательным звеном, который логично может замкнуть цепь рассуждений о волновых знаковых функциях генома. Нелокальность генома, как кодирующего и отчуждающего геноинформацию хромосомного континуума, уже заложена в его голографических функциях. Такой сорт информации распределен в геноме, как в голограмме и/или квазиголограмме, и как во фрактале одновременно. Это может иметь место, если рассматривать геном с чисто вещественных позиций. На таком уровне геноинформации еще не работает квантовая нелокальность. Геноголограмма, если "считывать" ее, приводит к тому, что вещество хромосом отчуждает знако-образные волновые фронты в качестве направляющих векторов биоморфогенеза.
Существенным, можно сказать, фундаментальным дополнением здесь мог бы выступать ЭПР-механизм. Как один из способов управления жизненными процессами, он придает принципиально новые потенции клеткам и тканям - возможность мгновенно передавать какие-то пулы генетико-метаболической информации между всеми клетками и тканями биосистемы, например, через поляризационный канал фотонов и радиоволн, о чем упоминалось выше. Если такой способ реален, тогда становится понятным, почему стратегические знаковые биомолекулы - нуклеиновые кислоты и белки имеют L-изомерный состав компонентов, спиральную закрутку и, соответственно, ярко выраженную способность к дисперсии оптического вращения, круговому дихроизму и двойному лучепреломлению. Теперь по-другому можно понимать и факт изомерной квантованности и других биоорганических молекул. Асимметричность их атомов и следующая отсюда изомерия и оптическая активность - это возможность быстрого автосканирования биосистемой поляризационной, голографической и иной вещественно-волновой информации о состоянии собственного метаболизма и о своей текущей сиюминутной тонкой пространственно-временной структуре.
Характерно, что успех экспериментальной квантовой телепортации достигнут, в частности, потому, что для генерации фотонов, разведения их в пространстве и их "программирования" использовали волноводы (световоды), лазеры с Уф накачкой и поляризаторы. Формально, перечисленные компоненты имеют биоаналоги в виде микротрубочек клеточного ядра и цитоплазмы, когерентных излучений ДНК и хромосом. Последние одновременно являются информационными биополяризаторами собственных лазерных излучений, а то, что ДНК и хромосомы являются лазероактивной средой показали наши прямые эксперименты [21].
Если ЭПР-фактор работает в биосистемах, логично спросить, почему организмы не ограничиваются столь эффективной формой оперирования биоинформацией и для чего им нужны еще и нервные импульсы, скорость прохождения которых (8-10м/сек.) весьма далека от скорости света в ДНК-квантовом биокомпьютере живых клеток? Можно высказать только предположение, что нервная система понадобилась высшим организмам, чтобы затормозить для переработки и усвоения слишком быстрых информационных процессов, к которым эволюция биосферы еще не подошла. Скорее всего, функции нервной системы и квантовой нелокальности генома комплементарны и сосуществуют, иногда давая всплески в виде паранормальных способностей людей-вычислителей, или в телепатии, не говоря уже о многих других "аномалиях" биосистем, частично теоретически осмысленных нами ранее [29, 30].
1.Maslov M.U., Gariaev P.P., 1994. Fractal Presentation of Natural Language Texts and Genetic Code. 2nd International Conference on Quantitative Linguistics "QUALICO-94", September 20-24, Lomonosov Moscow State University, Philological Faculty, pp. 193-194.
2.Gariaev P.P., Vasiliev A.A., Berezin A.A., 1994. Holographic associative memory and information transmission by solitary waves in biological systems. SPIE, The International Society for Optical Engineering, CIS Selected Papers, Coherent Measuring and Data Processing Methods and Devices, v. 1978, pp. 249-259.
3.Гаряев П.П., Внучкова В.А., Шелепина Г.А., Комиссаров Г.Г., 1994. Вербально-семантические модуляции резонансов Ферми-Паста-Улама как методология вхождения в командно-образный строй генома. Журнал русской физической мысли, N 1-4, с. 17-28.
4.Гаряев П.П., 1994, Кризис генетики и генетика кризиса. Русская мысль. М., Общественная польза, N 1-6, с. 46-49.
5.Трубников Б.А., Гаряев П.П., 1995, Похожа ли "речь" молекул ДНК на компьютерные программы? Природа, N1, с. 21 - 32.
6.Березин А.А., Гаряев П.П., 1995, Моделирование электроакустического излучения ДНК как носителя биоинформации., 2-й Международный симпозиум “Механизмы Действия Сверхмалых Доз Излучений”, 23-26 мая 1995г., Москва. с.122. (тезисы)
7.Гаряев П.П., Леонова Е.А., 1996, Генетический аппарат как волновая управляющая система. Международная научно-практическая конференция “Анализ систем на пороге XXI века: теория и практика”., с.69-78.
8.Готовский Ю.В., Комиссаров Г.Г., Гаряев П.П., 1996, Новая методика диагностики заболеваний по семи основным точкам акупунктуры (чакрам) и аппаратура для реализации. II Международная конференция “Теоретические и клинические аспекты биорезонансной и мультирезонансной терапии”. Центр Интеллектуальных Медицинских Систем “ИМЕДИС”. Москва, 1996г. с.164-169.
9.Щеглов В.А., Гаряев П.П., 1996, Лазер-лазерные взаимодействия и фантомные эффекты в генетических структурах. Материалы научной конференции с международным участием “Наука на пороге XXI века - новые парадигмы”. С.7.
10.Гаряев П.П., 1996, Генетический аппарат как волновая управляющая система. Международная научно-практическая конференция “Анализ систем на пороге 21 века. Москва. С.69-78.
11.Благодатских В.И., Гаряев П.П., Леонова Е.А., Маслов М.Ю., Шайтан К.В., Щеглов В.А., 1996, О динамике возникновения дислокаций в молекуле ДНК. Краткие сообщения по физике. Физический Институт РАН, N3-4, с.9-14.
12.Гаряев П.П., Маслов М.Ю., Решетняк С.А., Щеглов В.А., 1996, Взаимодействие электромагнитного излучения с информационными биомакромолекулами. "Антенная" модель. Краткие сообщения по физике. Физический Институт РАН, N1-2, с.54-59.
13.Гаряев П.П., Маслов М.Ю., Решетняк С.А., Щеглов В.А., 1996, Модель взаимодействия электромагнитного излучения с информационными биомакромолекулами., Краткие сообщения по физике. Физический Институт РАН, N1-2, с.60-63.
14.Гаряев П.П., Маслов М.Ю., Решетняк С.А., Щеглов В.А. 1996, Модель взаимодействия электромагнитного излучения с информационными биомакромолекулами. Краткие сообщения по физике. Физический Институт РАН, N1-2, с.60-63.
15.П.П.Гаряев, М.Р.Гарбер, Е.А.Леонова., 1998, Виртуальный геном прионов. Фридмановские чтения. Всероссийская научная конференция. г.Пермь, 7-12 сентября 1998г. с.140-142.
16.Гаряев П.П., Тертышный Г.Г., Готовский Ю.В., 1997, Трансформация света в радиоволны. III международная конференция “Теоретические и клинические аспекты применения адаптивной резонансной и мультирезонансной терапии”. “ИМЕДИС”. Москва. 18-20 апреля 1997г. с.303-313.
17.Гаряев П.П., Леонова Е.А., 1996, Пересмотр модели генетического кода. Сознание и Физическая Реальность. Изд. "ФОЛИУМ", т.1, N1-2, с.73-84.
18.S.A.Reshetnyak, V.A.Shcheglov, V.I.Blagodatskikh, P.P.Gariaev, and M.Yu.Maslov, 1996, Mechanism of interaction of electromagnetic radiation with a biosystem, Laser Physics, v.6, N2, pp.621-653.
19.Berezin A.A., Gariaev P.P., Gorelik V.S., Reshetniak S.A., Shcheglov V.A., 1996, Is it possible to create laser based on information biomacromolecules? Laser Physics, v.6, N6, pp.1211-1213.
20.A.A.Berezin, P.P.Gariaev, S.A.Reshetniak, K.V.Shaitan, V.A.Shcheglov, 1996, To the problem of possible development at a biolaser working on Frolich modes. (препринт Физического Института им.П.Н.Лебедева №49, 12с.)
21.А.М.Агальцов, П.П.Гаряев, В.С.Горелик, И.А.Рахматуллаев, В.А.Щеглов, 1996, Двухфотонно-возбуждаемая люминесценция в генетических структурах. Квантовая электроника, v.23, N2, с.181-184.
22.П.П.Гаряев, 1996, Эпигенетическая роль внеклеточных матриксов. Гипотеза кодовой иерархии. Межреспубликанский заочный научно-технический семинар “Применение лазеров в науке и технике”, вып.8. Иркутск. Изд. Иркутского Филиала Института Лазерной Физики СО РАН, с.85-107.
23.П.П.Гаряев, 1996, Информационно-волновые свойства живых систем. Голографический аспект. Межреспубликанский заочный научно-технический семинар “Применение лазеров в науке и технике”, вып.8. Иркутск. Изд. Иркутского Филиала Института Лазерной Физики СО РАН, с.137-159.
24.П.П.Гаряев, 1996, О природе рефлексотерапии. Современные концепции первичных механизмов акупунктуры и акупрессуры. Межреспубликанский заочный научно-технический семинар “Применение лазеров в науке и технике”, вып.8. Иркутск. Изд. Иркутского Филиала Института Лазерной Физики СО РАН, с.188-206.
25.Гаряев П.П., Леонова Е.А., 1996, Новая модель генетического кода. Сборник научных трудов. Академия медико-технических наук РФ. Отделение “Биотехнические системы и образование” при МГТУ им. Н.Э.Баумана. Выпуск 1. с.25-34.
26.Гаряев П.П., Тертышный Г.Г., 1997, Явление перехода света в радиоволны применительно к биосистемам. Сборник научных трудов. Академия медико-технических наук РФ. Отделение “Биотехнические системы и образование” при МГТУ им. Н.Э.Баумана. Выпуск 2. с. 31-42.
27.Гаряев П.П., Тертышный Г.Г., Лощилов В.И., Щеглов В.А., Готовский Ю.В., 1997, Переход лазерного света в электромагнитное излучение радиодиапазона Сборник научных трудов. Академия медико-технических наук РФ. Отделение “Биотехнические системы и образование” при МГТУ им. Н.Э.Баумана. Выпуск 2. с.31.
28.Прангишвили И.В., Гаряев П.П., Тертышный Г.Г., Максименко В.В., Мологин А.В., Леонова Е.А., Мулдашев Э.Р., 2000, Радиоволновая спектроскопия локализованных фотонов: выход на квантово нелокальные биоинформационные процессы. Датчики и системы, №9 (18), с.2-13.
29.Гаряев П.П., 1994, Волновой геном. Российская Академия Наук. Изд. Общ. Польза. Москва. 279с.
30.Гаряев П.П., 1997, Волновой генетический код. Издатцентр. Москва. 108с.
31.Mantegna R.N., Buldyrev S.V., Goldberger A.L., Havlin S., Peng S.-K., Simons M. and Stanley H.E. Linguistic Features of Noncoding DNA Sequences.
// Phys.Rev.Lett. 1994.v.73. № 23. p.3169- 3172.
32.П.П.Гаряев, М.Р.Гарбер, Е.А.Леонова, Г.Г.Тертышный, 1999, К вопросу о центральной догме молекулярной биологии. Сознание и физическая реальность, Изд. "ФОЛИУМ", т.4, №1, с.34-46.
33.P.Gariaev, G.Tertishniy, 1999, The quantum nonlocality of genomes as a main factor of the morphogenesis of biosystems. III Sci. and Medical Network Continental Members Meeting. Schlob Petzow near Potsdam, Germany, May 6-9, 1999. p.37-39.
34.Приоритет на патент по международной заявке на изобретение №99/01/Л от 06.01.1999.
35.Maksimenko V.V., 1999, Localization of Photon between Pair of Particles.-1. Elastic Scattering, Journal of Aerosol Science, v. 30, p. 287.
36.Maksimenko V.V., 1999, Localization of Photon between Pair of Particles.-2. Inelastic Scattering, Journal of Aerosol Science, v. 30, p. 289.
37.Maksimenko V.V., 1999, Localization of Light in Fractal Cluster, Journal of Aerosol Science, v. 30, p. 291.
38.Гаряев П.П., Тертышный Г.Г., Готовский Ю.В., Леонова Е.А., 1999, Голографическая и квантовая нелокальность генома. // 5-я междунар. Конф. "Теорет. и клинич. аспекты применения биорезонансной и мультирезонансной терапии", часть II. "Имедис". Москва, 1999г. с.256-272.
39.J.A.Tuszinski, R.Paul, Chatterjee R, Sreenivasan S.R., 1984, Relationship between Frolich and Davidov models of biological order. Physical Review A, v.30, N5, pp.2666-2675.
40.Давыдов А.С., 1984, Солитоны в молекулярных системах. Киев. Наукова Думка. 288с.
41.Frolich H., 1968, Phys. Lett., v.26A, p.402.
42.Frolich H., 1972, Phys. Lett., v.29A, pp.153-154.
43.Frolich H., 1975, Proc.Natl.Acad.Sci., USA, v.72, pp.4211-4215.
44.Frolich H., 1977, Neurosci.Res.Programm.Bull., v.15, pp.67-72.
45.В.И.Благодатских, П.П.Гаряев, М.Ю.Маслов, С.А.Решетняк, В.А.Щеглов, 1996, Взаимодействие когерентного электромагнитного излучения с биомакромолекулами, ч. 1, Laser Physics, v.6, N4, pp.621-653.
46.И.В.Прангишвили, П.П.Гаряев, Г.Г.Тертышный, Е.А.Леонова, А.В.Мологин, М.Р.Гарбер, 2000, Генетические структуры как источник и приемник голографической информации. Датчики и системы, №2(11), с.2-8.
47.Мулдашев Э.Р., 2000, Комбинированная трансплантация глаза. Мин. здрав. РФ. Всеросссийский центр глазной и пластической хирургии. “Аллоплант”. 30с. Препринт.
48.Прангишвили И.В., Гаряев П.П., Тертышный Г.Г., Мологин А.В., Леонова Е.А., Мулдашев Э.Р., 2000, Голографическая модель 3-мерного биоморфогенкеза. Датчики и системы (в печати).
49.Adleman L., 1994, Molecular computation of solutions to combinatorial problems. Science, v.266, pp.1021- 1024.
50. Цит. по статье “Gene Genie”. T.A.Bass, WIRED Archive 3.08. Aug. 1995, 8p. Интернет. Сайт <Leonard Adleman>.
51.Без учета наших моделей кодирования информации в ДНК.
52.Gariaev P.P., 1994, DNA as source of new kind of God "knowledge", Act and Facts/Impact series, N12, pp,7-11.
53.Einstein A., Podolsky B., Rosen N., 1935, Can Quantum-Mechanical Description of Physical Reallity be Considered Complete? Phys.Rev. v.47, p.777-780.
54.Bell J.S., 1964, On the Einstein-Podolsky-Rosen Paradox, Physics, v.1, pp.195-200; Bell J.S., 1976, Einstein-Podolsky-Rosen Experiments. Proceed. Symp. On Frontier Problems in High Energy Physics (Pisa), p.33-45.
55. Bennet C.H., Brassard G., Crepeau C., Jossa R., Peres A., Wootters W.K., 1993, Teleporting and unknown quantum state via dual classical and Einstein-Podolsky-Rosen channels. Phys.Rev.Lett.,v.70, p.1895-1899.
56. Bouwmeester D., Pan Jian-Wei, Mattle K., Eibl M., Weinfurter H., Zeilinger A., 1997, Experimental quantum teleportation. Nature, v.390, p.575-579.
57. Josephson B.D., Pallikari-Viras F., 1991, Biological Utilisation of Quantum NonLocality. Foundation of Physics, v.21, p.197-207, Plenum Press ©.
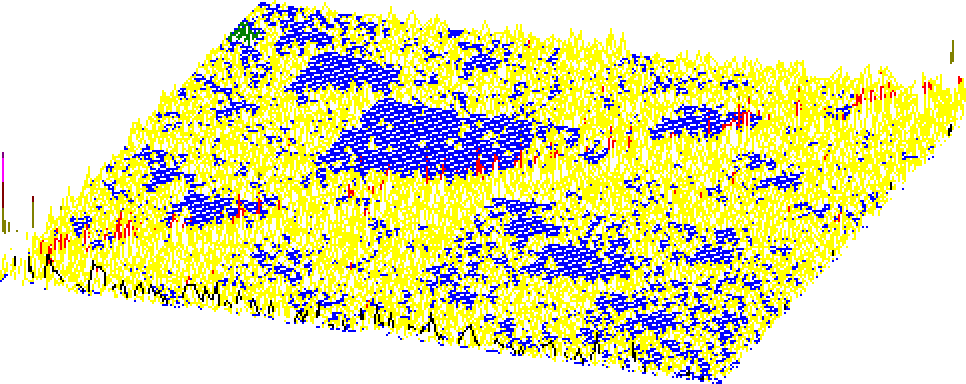
Рис.1б) Матрица плотности хаотически-игрового представления текста на английском языке (Шилд Б., 1992, Руководство по компьютерному программированию. 250с). Анализировали последовательность появления в тексте четырёх слов. Левый ближний угол соответствует слову "the", правый ближний - слову "in", левый дальний - "on", правый дальний - "of".

Рис.2а) Влияние нуклеотидной последовательности ДНК на динамику конформационного возмущения уединённой (солитоноподбной ) волны. Последовательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). центр возмущения - 400-ый нуклеотид.

Рис. 2.б) То же, что Рис.2 а, но центр возмущения цепочки ДНК на 450-ом нуклеотиде.
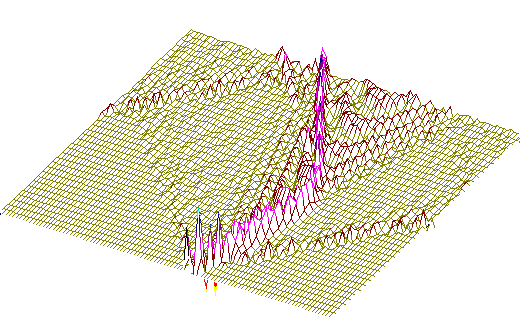
Рис. 3.а) Та же модель, что на рис.2, но с учётом нелинейности ковалентных связей в сахаро-фосфатном остове ДНК.Последовательность нуклеотидов - вирус саркомы птиц (первые 600 пар оснований). Центр возмущения - 200-ый нуклеотид.
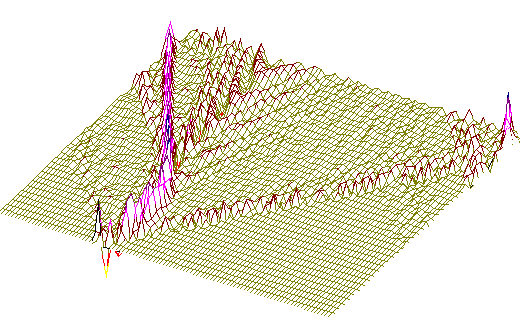
Рис. 3.б) То же, что на Рис 2, но центр возмущения - 400-ый нуклеотид.

Рис. 3.в) То же, что на Рис 2, но центр возмущения - 500-ый нуклеотид.
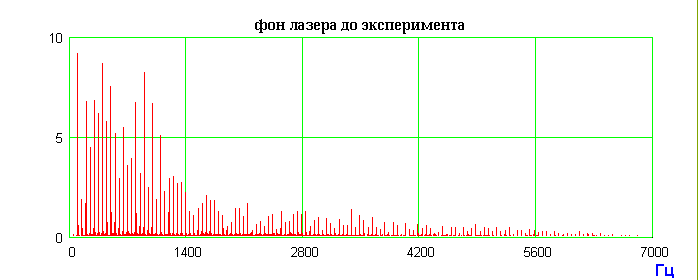
Радиоволновой фон лазера до эксперимента
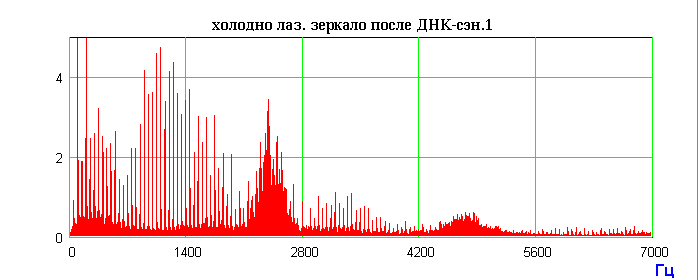
Радиоволновой сигнал образца ДНК, записываемый и генерируемый внутренними горячими лазерными зеркалами.
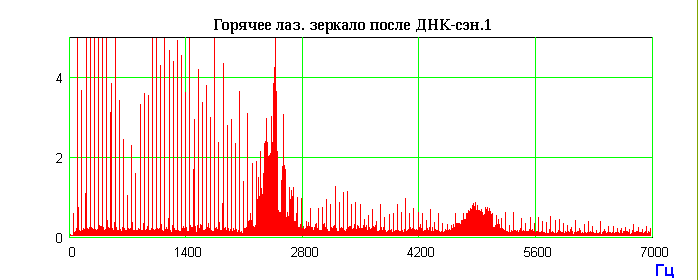
Запись радиосигнала на внешнем холодном лазерном зеркале, которое при комнатной температуре помещали позади рабочего лазера в режиме прохождения луча через образец ДНК и отражения луча в резонатор лазера.
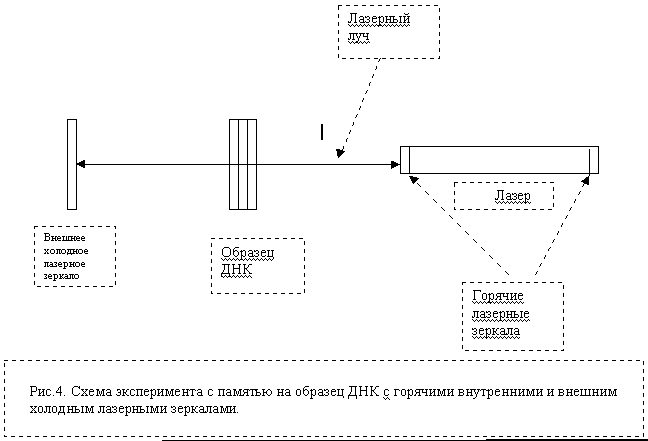
Рис.4. Схема эксперимента с генерацией индуцированных радиоволновых сигналов препаратом ДНК и запись радиоволновых сигналов с внутреннего и внешнего лазерных зеркал. Последовательность операций: фиксировали радиоволновой фон лазера до экспериментов; после экспонирования лазера и внешнего холодного зеркала препаратом ДНК и фиксации индуцированного радиоволнового спектра ДНК препарат ДНК и внешнее зеркало убирали; через 3 часа радиоволновое излучение лазера достигало исходного фонового уровня; после этого внешнее холодное зеркало устанавливали на прежнее место в режиме отражения луча в резонатор лазера; фиксировали радиоволновой спектр, аналогичный индуцированному спектру препарата ДНК; внешнее холодное лазерное зеркало хранит запись спектра препарата ДНК около 3 месяцев.
Главная страница Список все работ и книг
Прислано в феврале 2001.